Viagra gibt es mittlerweile nicht nur als Original, sondern auch in Form von Generika. Diese enthalten denselben Wirkstoff Sildenafil. Patienten suchen deshalb nach viagra generika schweiz, um ein günstigeres Präparat zu finden. Unterschiede bestehen oft nur in Verpackung und Preis.
Oxidative stress and microcirculatory flow abnormalities in the ventricles during atrial fibrillation
*, , Christopher H. Lillig 3 and
1 Department of Cardiology and Intensive Care Medicine, St. Vincenz-Hospital Paderborn, Paderborn, Germany2 Working Group: Molecular Electrophysiology, University Hospital Magdeburg, Magdeburg, Germany3 Institute of Medical Biochemistry and Molecular Biology, Ernst-Moritz-Arndt University, Greifswald, Germany
Patients with atrial fibrillation (AF) often present with typical angina pectoris and mildly
David R. Van Wagoner, Cleveland
elevated levels of cardiac troponin (non-ST-segment elevation myocardial infarction) during
Clinic Lerner College of Medicine of
an acute episode of AF. However, in a large proportion of these patients, significant coro-
Case Western Reserve University,USA
nary artery disease is excluded by coronary angiography, which suggests that AF itself
influences myocardial blood flow. The present review summarizes the effect of AF on the
Barbara Casadei, University of
occurrence of ventricular oxidative stress, redox-sensitive signaling pathways and gene
Oxford, UK
expression, and microcirculatory flow abnormalities in the left ventricle.
David R. Van Wagoner, ClevelandClinic Lerner College of Medicine of
Keywords: angiotensin, atrial fibrillation, microvascular flow, oxidative stress
Case Western Reserve University,USAGeorge John Rozanski, University ofNebraska Medical Center, USA
*Correspondence:
Andreas Goette, Department of
Cardiology and Intensive Care
Medicine, St. Vincenz-Hospital
Paderborn, Am Busdorf 2, 33098
Paderborn, Germany.
e-mail:
VENTRICULAR OXIDATIVE STRESS AND SIGNAL
Angina pectoris is a typical symptom in patients with parox-
TRANSDUCTION DURING AF
ysmal atrial fibrillation (AF). In most of these patients, angina
Reactive oxygen species (ROS) are generated under physiologi-
pectoris is associated with mildly elevated cardiac troponin (cTn)
cal conditions in the cardiovascular system and act as second
levels suggesting a non-ST-segment elevation myocardial infarc-
messengers in numerous redox-sensitive signal transduction path-
tion (NSTEMI). However, in a large proportion of these patients,
ways However, under pathophysiological conditions,
significant coronary artery disease can be excluded by coronary
chronically elevated amounts of ROS may exert oxidative stress.
angiography despite clinical symptoms
Historically, the term "oxidative stress" was defined as an imbal-
Although the elevated ventricular rate during AF
ance between the generation of ROS and the capacity of the defense
may contribute to the symptoms of angina pectoris
systems During the past decade, this model
angina pectoris develops also in patients with a slow ven-
has evolved based on some key findings: the production of dif-
tricular rate and most patients tolerate fast ventricular rates in
ferent oxidants affects distinct presets of target proteins through
sinus rhythm without any clinical symptoms
modifications that are specific both with respect to the oxidant
Recent reports suggest that myocar-
and the site of modification, most frequently well-defined cys-
dial blood flow is reduced, whereas coronary vascular resistance
teinyl side chains. The so-called antioxidant redox systems in the
is elevated in patients with AF
different cellular compartments, e.g., glutathione, NADPH, thiore-
One potential link between AF, abnormal ventric-
doxin (Trx), and peroxidases such as the peroxiredoxins (Prx), are,
ular perfusion, and cardiomyocyte dysfunction is the occurrence
however, not in equilibrium and independently maintained at dis-
of oxidative stress and the disruption of redox signaling through
tinct redox potentials. Oxidative stress may thus, more timely, be
activation of the nicotinamide adenine dinucleotide phosphate
defined as the chronic disturbance of redox circuits and redox-
oxidase (NADPH oxidase;
responsive signal transduction pathways
Repetitive episodes of AF-induced
ventricular ischemia may contribute to the development of a
Oxidative stress has been implicated as playing a critical
pathological vicious cycle combining AF and left ventricular (LV)
role in the pathophysiology of heart and cardiovascular dis-
eases such as heart failure, LV hypertrophy, coronary heartdisease,
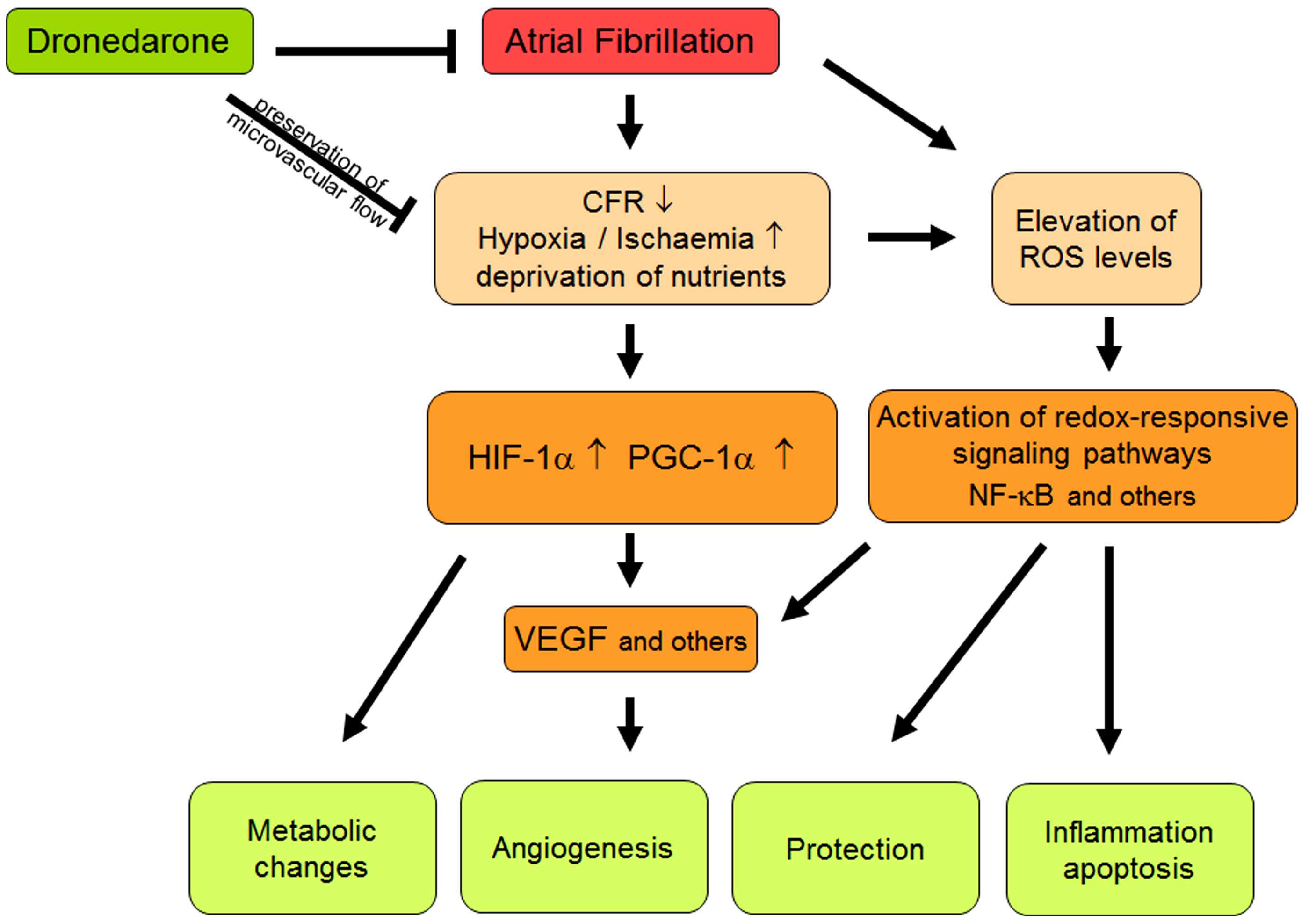
Goette et al.
Ventricular oxidative stress during AF
FIGURE 1 Schematic summary of different interactions between atrial fibrillation, oxidative stress, and flow abnormalities. Abbreviations should be
included here are explained in the text.
cardiac arrhythmia. It appears that the oxidative events initiate
as reoxygenation-induced PV burst firing represent important
the disease-dependent tissue remodeling and promote its prop-
proarrhythmogenic mechanisms
agation. Oxidative stress is associated with microvascular flow
Coronary flow can be estimated in vivo using wire-based sys-
abnormalities and occurs immediately after new-onset AF likely
tems Coronary flow
representing key initiator mechanisms of AF-related ventricular
reserve (CFR) measurements can be measured using a pressure
remodeling. This has been shown in patients with lone recur-
temperature sensor-tipped guidewires, which allow the simulta-
rent AF and for rapid atrial pacing models
neous determination of the fractional flow reserve (FFR). CFR
measurements are influenced by flow abnormalities in the epicar-
ular RR intervals are considered to be responsible for compro-
dial arteries and the microcirculation. In contrast, reduced FFR is
mised coronary blood flow reserve, thus preventing the adequate
specific to epicardial lesions. Recent studies can clearly show that
attainment of the increased oxygen demand due to tachycardia
acute episodes of AF induce oxidative stress in the LV myocardium
In addition, increased catecholamine
and compromise microvascular blood flow
levels via α-adrenergic vasoconstriction may further contribute
to AF-induced ischemia AF-
Although it is well established that ischemia creates a sub-
dependent cardiac remodeling, especially fibrosis, may well con-
strate for AF maintenance
tribute to long-term restriction of microcirculation
the effects of AF on ventricular microcirculation and underlying
Resulting imbalances of myocardial oxygen supply and
pathways are less well understood. Impaired intracellular Ca2+-
myocardial oxygen demand lead to the specific activation of redox-
handling as described above together with elevated cardiac and
sensitive signaling pathways, that are either protective or part
systemic angiotensin II levels are two important factors which very
of the pathophysiological process causing onset and progression
likely contribute to the AF-dependent activation of redox-sensitive
signaling pathways in the ventricles. These will be discussed in
Ischemia facilitates the onset of AF by altering cellular ionic
more detail below.
homeostasis, in particular via tachycardia-induced intracellularcalcium and sodium overload. Increased spontaneous ectopy is
VASOCONSTRICTORY PEPTIDES REGULATING CORONARY FLOW
due to increased NCX currents and spontaneous Ca2+-release
events In pulmonary veins (PV), hypoxia-
Vasoactive peptides such as angiotensin II (AngII) and endothe-
induced EAD, and delayed after-depolarizations (DAD) as well
lin (ET) play important roles in the regulation of cardiovascular
Goette et al.
Ventricular oxidative stress during AF
function as well as in the pathogenesis of heart and cardiovascular
of hypercholesterolemic animals In a recent
diseases. AF has been associated with increased plasma and tissue
study it was shown that application of irbesartan prevents ventric-
levels of angiotensin II (AngII)
ular oxidative stress and microvascular flow abnormalities during
which are resulting from increased expression/activity of
7 h of AF Nevertheless, clinical trials like the
ACTIVE study failed to demonstrate a measurable benefit of long-
Angiotensin II mediates its major hemodynamic and patho-
term ARB therapy in patients with AF
physiological effects via the AT1 receptor. Six hours of rapid atrial
pacing is sufficient to elevate plasma AngII levels suggesting thevery early involvement of this vasoactive molecule in the patho-
genesis of AF The activation of the AT1
Endothelin-1 (ET-1) exerts its proarrhythmogenic effects by two
receptor induces a cascade of phosphorylation events that even-
different ways. First, due to its strong vasoconstrictory activity,
tually activates MAP kinases, which stimulate the proliferation
ET-1 may induce ischemia which facilitates arrhythmia. Second,
of fibroblasts, cellular hypertrophy, and apoptosis. Furthermore,
upon binding to endothelin receptor A (ET-A), ET-1 affects intra-
activation via the AT1 receptor releases calcium from intracellu-
cellular calcium handling and, in particular, provokes intracellular
lar stores and activates protein kinase C (PKC). PKC regulates
Ca2+-waves via IP3-dependent Ca2+-release leading to DAD
the function of calcium and potassium channels, and phosphory-
Furthermore, ET-1 activates PKC and MAP kinases
lates p47phox regulatory subunit. The phosphorylation of p47phox
Atrial stretch is a potent factor promoting the
plays a pivotal role in the activation of NOX2/NOX1 by providing
production and release of ET-1 ET-1 acti-
physical binding domains to another regulatory subunit p67phox
vates NADPH oxidase via the ET receptor-proline-rich tyrosine
It is noteworthy that both the MAP kinase
kinase-2 (Pyk2)-rac1 pathway
as well as the calcium/PKC signaling pathways respond to ROS
The GTPase, rac1, binds to p67phox and activates NADPH oxidase
themselves and are regulated by redox effector molecules from the
in its GTP-bound state
Trx family of proteins
valvular disease
At the atrial level, it is well established that AngII upon bind-
primary pulmonal hypertension but
ing to its preferred receptor, AT1R, leads to the activation of
also AF are all associated with increased plasma and cardiac tissue
levels of ET-1. Atrial ET-1 levels were correlated with atrial rhythm,
In blood vessels, AngII infusion resulted in increased
atrial size, and hypertension and were associated with hypertro-
expression and activity of NADPH oxidase, which was dependent
phy, fibrosis, and atrial dilatation Changes
on PKC to some extent In the murine heart,
in cardiac and circulating levels of ET-1 have been described in
AngII increased superoxide generation and cardiac hypertrophy
experimental models of myocardial ischemia and in patients with
involving NOX2 Increased NADPH oxidase
acute myocardial infarction
activity contributes to elevated ROS production, protein modi-
Accordingly, a dual ET receptor antago-
fication, and redox-related gene expression patterns observed in
nist has been demonstrated to prevent coronary vasoconstriction
AF Moreover, it has been identified as an
during reperfusion of ischemic heart AF-
independent risk factor for post-operative AF
dependent negative changes in the microcirculation
Pre-operative treatment with ascorbate prevented AF to a signifi-
contribute to and further enhance
cant extent Similarly, ascorbate reduced the
increased gene expression and release of ET-1 which, in turn,
recurrence of AF after cardioversion
aggravates coronary vasoconstriction
Less is known about the effects of AF on ventricular func-
and leads to oxidative stress
tion and underlying changes in redox-signaling pathway activity
Although the AngII/ET-1-induced increase of ROS generation is
and related gene expression. AF is associated with increased coro-
mostly associated with hemodynamic response and development
nary resistance, compromised vasodilation
of hypertension, solid evidence shows that these vasoactive pep-
reduced coronary flow or flow reserve
tides via activation of NADPH oxidase mediate changes in vascular
As these changes can be largely
architecture and heart damage
attenuated by the administration of sartans
AngII seems to be a major factor linking AF with LV malperfusionand dysfunction. In support of this view, the RAP-induced Ang-II-
dependent activation of NADPH oxidase and down-stream effec-
It has been suggested that the NADPH oxidase is an important
tors of redox-activated signaling cascades, e.g., LOX-1 expression
source of ROS in the left ventricle during atrial tachyarrhyth-
and F2-isoprostane formation, were all attenuated by irbesartan
mia NADPH oxidase
was originally discovered in neutrophils, where, during phago-
At the molecular level, ANG-II-receptor blockers (ARBs) have
cytosis, millimolar quantities of superoxide can be released into
been shown to increase NO availability and it is possible that this
the extracellular (phagosomal) compartment. In non-phagocytic
effect is mediated by a stronger ANG-II-dependent activation of
cells such as cardiomyocytes, fibroblasts, and endothelial cells,
AT2-receptors. ARBs also attenuate aortic intimal proliferation
the amounts of produced superoxide are much lower and occur
and markedly decrease the enhanced LOX-1 expression in the aorta
mostly intracellularly The neutrophil NADPH
Goette et al.
Ventricular oxidative stress during AF
oxidase is composed of membrane-associated subunits: p22phox,
ROS production is shifted from NADPH oxidase to mitochon-
and NOX, and four cytosolic regulatory subunits: p47phox, p67phox,
drial oxidases and uncoupled eNOS in the right atrium
p40phox, and the small GTPase rac1 or rac2. NOX is the key cat-
This is in full accordance with the observation that
alytic subunit of the NADPH oxidase and in non-phagocytic cells
statins, which reduce ROS production by NADPH oxidases via
possess several isoforms. Beside the phagocyte NADPH oxidase
inhibition of Rac1, are effective in acute models of AF and in
(NOX2; gp91phox), the expression of six homologs (NOX1, NOX3,
patients with post-operative AF, but fail to reduce ROS production
NOX4, NOX5, Duox1, and Duox2) has been identified
in models of long-lasting AF or patients with permanent AF. Pre-
While the cytosolic regulatory components translocate to
operative statin-treatment was shown to reduce myocardial O −
the membrane to form the active NADPH oxidase complex upon
and ONOO− production by reducing NADPH oxidase activity
activation in the neutrophil in the non-
phagocytic cells preassembled functional enzyme complex is partlypresent intracellularly
VENTRICULAR MICROCIRCULATION DURING AF
Although the non-phagocyte NADPH oxidase is consti-
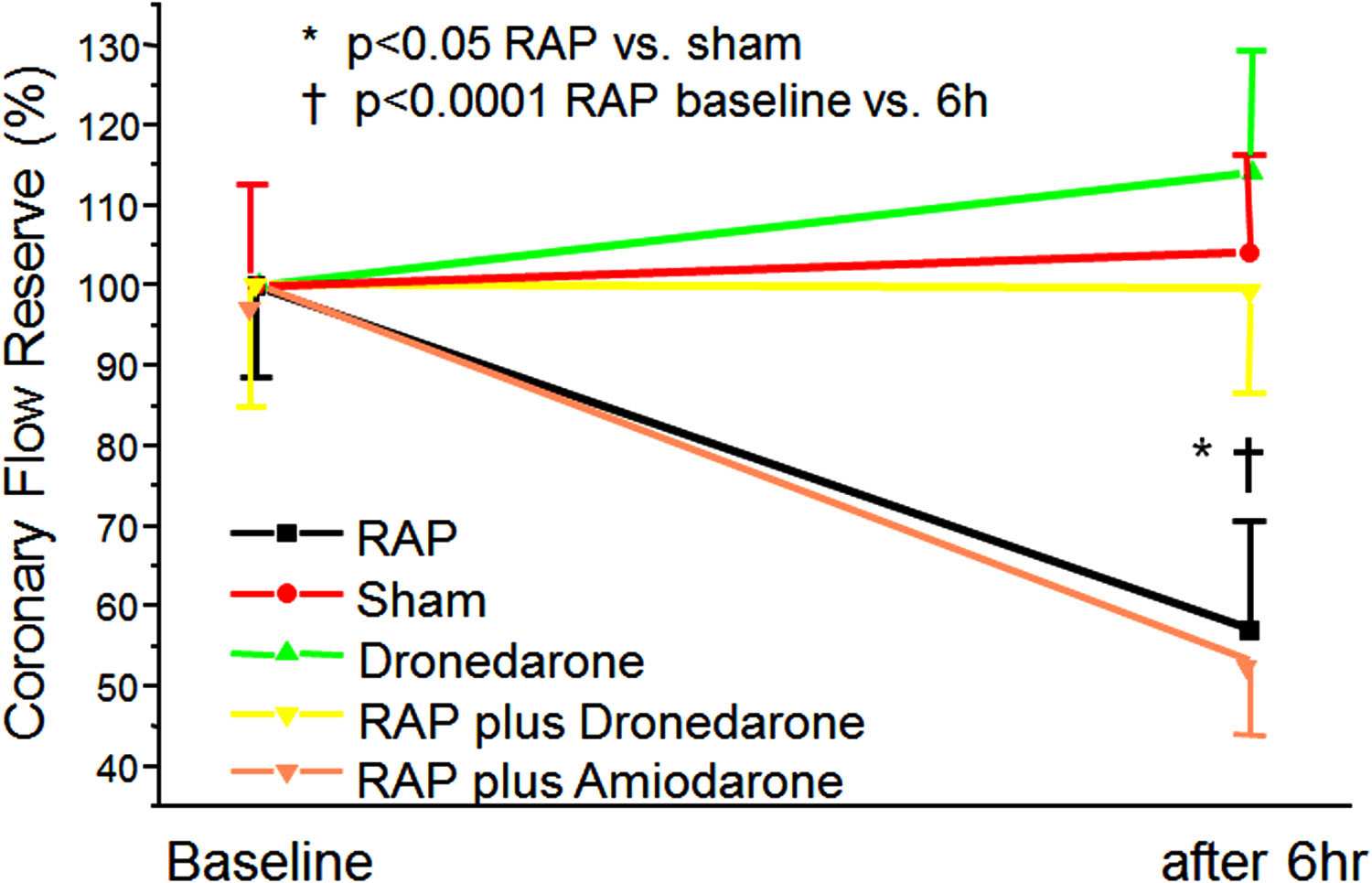
An induced episode of AF of up to 6 h has no effect on FFR (marker
tutively active, its activity can be further up-regulated in response
for epicardial flow) in pigs In contrast,
to vasoactive peptides (AngII, ET-1), hormones, growth factors,
CFR (index of microvascular abnormalities if FFR is normal) is
cytokines, and mechanical stress
substantially reduced (about 50%) after an AF episode of 6 h. Inter-
estingly, application of irbesartan and dronedarone could prevent
The NOX-dependent ROS signaling is an important factor
microcirculatory flow abnormalities to occur whereas amiodarone
responsible for development of many pathological processes in
has no effect on CRF These results correspond to latest
the ventricles during cardiac hypertrophy, remodeling, and heart
findings that dronedarone reduces the size and volume of induced
failure Recently, in an animal model of
cerebral and myocardial infarcts
acute AF, increased expression of NOX2, NOX1, and enhanced
Patients without previously documented coro-
expression of NOX4 were shown in the left ventricle after 6 h of
nary artery disease sometimes develop chest discomfort with the
atrial pacing The AT1
onset of AF Furthermore, patients with AF
receptor antagonist, irbesartan, and the multichannel inhibitor,
have ventricular-flow abnormalities and a higher incidence of car-
dronedarone, efficiently prevented the up-regulation of NOX2
diac events Consistent
The elevated expres-
with this notion, coronary artery resistance is markedly elevated
sion of NOX2 and superoxide production in the left ventricle was
(by 62%), whereas myocardial blood flow is substantially reduced
also observed in a rabbit model of chronic heart failure
in AF patients Vasodilatation in response
Several studies have shown a crucial role of NOX2
to exercise is also compromised during AF
in the response to AngII-induced LV hypertrophy
The Doppler-derived coronary vascular resistance index has been
It was found that the regu-
reported to be increased by 67% in an experimental AF model
latory compound rac1 initiated hypertrophic response
Induction of AF for up to 6 h has no effect on
Amounts of the regulatory p47phox
FFR (marker of epicardial flow) in pigs
increase after myocardial
by contrast, CFR (index of microvascular abnormalities if FFR
infarction in the left ventricle and contribute to the NADPH oxi-
is normal) is substantially reduced (about 50%) by short-term
dase dependent tissue remodeling NOX2
AF. Interestingly, irbesartan and dronedarone could prevent the
and NOX4 are the main isoforms expressed in the cardiac cells.
occurrence of microcirculatory flow abnormalities whereas amio-
NOX4, in contrast to NOX2, does not require the presence of
darone had no effect These results are in keeping with
regulatory oxidase proteins p47phox or the GTPase rac. Moreover,
recent findings indicating that dronedarone reduces the size and
NOX4 produces mainly hydrogen peroxide and only very small
volume of induced cerebral and myocardial infarcts
amounts of superoxide intracellularly and
was found on internal membranes, in mitochondria and also in perinuclear endoplasmic
AF ALTERS VENTRICULAR EXPRESSION OF
reticulum The up-regulation in NOX4 expres-
ISCHEMIA/HYPOXIA-RELATED GENE PANELS
sion was accompanied by mitochondrial dysfunction and apop-
Atrial fibrillation provokes rapid and profound changes in the ven-
tosis in the cardiomyocytes
tricular expression of ischemia/hypoxia-related genes
NOX1 is an important isoform expressed particularly in vascular
These expression changes were associated with and
smooth muscle cells and is responsible for extracellular superox-
may result partially from microcirculatory abnormalities. Both
ide production in coronary arterial myocytes. Several studies have
the observed RAP-dependent limitation of flow reserve and the
provided evidence that NOX1 oxidase is involved in mediating the
expression changes could be prevented by dronedarone. Interest-
hypertensive response to AngII in particular
ingly, amiodarone does not reduce AF-induced flow abnormal-
ities in the microvascular tree The positive effect of
Recent data suggests, however, that induction of atrial NADPH
dronedarone in brief episodes of AF is supported by the ATHENA
oxidase activity or subunit expression is an early but transient
trial, which found a reduced rate of acute coronary syndromes and
mechanism in the natural course of AF development and pro-
reduced cardiovascular mortality in patients with AF
gression With increasing duration of AF,
Goette et al.
Ventricular oxidative stress during AF
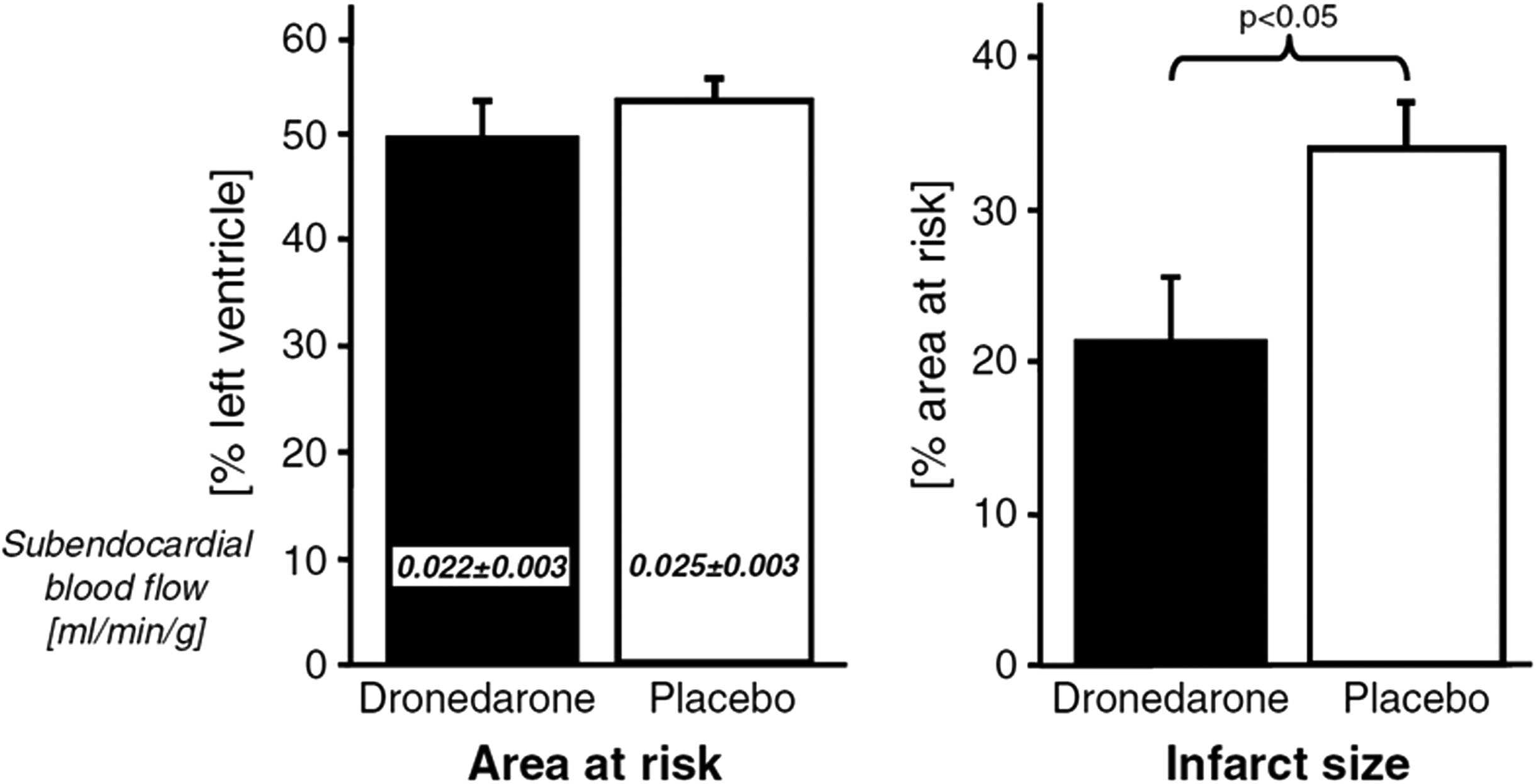
A recent study could show that acute application of
were HIF-1, VEGFA, and PPARGC1α, all of them known to be
dronedarone during an induced myocardial infarction reduced
induced in response to hypoxia or deprivation of nutrients
the infarct size substantially Nevertheless, in long-
The regulation of VEGF expression in response
lasting (6 month) AF, myocardial NADPH is not activated
to hypoxia is mediated by HIF-1α Under
and thereby, positive effects through inhibition of
the same conditions, and independent of this canonical HIF-
NADPH are unrealistic to exist. Thus, the antioxidant effects of
pathway, increased PPARGC1α exerts its strong angiogenic activity
dronedarone should not be present in permanent AF. This is sup-
and induces VEGF expression by co-activating ERR-α
ported by the PALLAS trial, which showed negative outcome if
Thus, both HIF-1α and PPARGC1α appear to be criti-
patients with permanent AF are treated with dronedarone. In PAL-
cally involved in the angiogenic response to AF-dependent flow
LAS, rates of stroke, myocardial infarction, and heart failure were
alterations and may provide protection against ischemic damage.
almost doubled in dronedarone treated patients
Rapid atrial pacing also led to an activation of the NF-κB
Thus, the therapeutic effect of dronedarone depends on the
pathway in the left ventricle ROS as intra-
duration of AF, which is quite a unique finding.
cellular messengers and redox effector molecules such as Trx and
Transcriptome analyses provided a first mechanistic insight
glutaredoxin (Grx) lead to the activation and nuclear translocation
into the pathophysiological processes mediating or even counter-
of this redox-sensitive transcription factor
acting coronary and ventricular dysfunction during AF. Among
Consistent with the RAP-dependent ventricular activation
the genes the expression of which was changed in response to RAP
of NF-κB, the expression of a panel of established down-streamtargets of NF-κB including VEGFA Fn14, CCL2 HIF1A as well as DnaJ fam-ily members, DNAJA4 and DNAJB9, that have been describedas co-chaperones for the ATPase activity of Hsp70 and func-tion to protect stressed cells from apoptosis was up-regulated in the left ventricle Both DNAJA4 and DNAJB9, but also thioredoxin (Trx1; and peroxiredoxin I (PrxI; are antioxidant response element (AREs) regulated genes acti-vated through nuclear factor-erythroid 2-related factor 2 (Nrf2) inresponse to oxidative stress. After phosphorylation by, e.g., PKC,Nrf2 translocates to the nucleus where it binds to AREs and trans-activates target genes of, e.g., enzymes such as PrxI that regulate theintracellular amounts of ROS It seems
FIGURE 2 Induction of ventricular-flow abnormalities in the ventricles
reasonable to assume that increased expression of anti-oxidative
during rapid atrial pacing (RAP) in comparison to unpaced controls
response genes, e.g., peroxiredoxins and DnaJ family members, is
using a porcine model. Effects of dronedarone and amiodarone
aimed at limiting stress-mediated tissue-damage. In this in vivo
demonstrated. Abbreviations are explained in the text adopted from
model of acute AF, dronedarone attenuated most of the ven-
tricular changes in gene expression. In addition, RAP-dependent
FIGURE 3 Effect of dronedarone on size of acute myocardial infarctions adopted from .
Goette et al.
Ventricular oxidative stress during AF
PKC phosphorylation, NADPH isoform expression, isoprostane
generally mediate the shift from aerobic metabolism and fatty acid
release, and IκBα phosphorylation were decreased. This, together
utilization to glucose utilization via pyruvate oxidation or even
with the attenuation of negative flow alterations may indicate that
to glycolytic metabolism
dronedarone beneficially affects very early steps of RAP-associated
HIF-1, together with c-myc, mediates the induction of HK2
ventricular pathology, very likely by preventing ischemia/hypoxia
which contributes to shift glucose away from mitochondrial uti-
lization and has also anti-oxidative effects
PPARGC1, the multi-functional co-activator, is also involved
On this background, the observed ventricular induc-
in the regulation of cardiac mitochondrial functional capacity
tion of glycolytic gene expression during AF completely fits to the
and cellular energy metabolism. In accordance with the observed
increased expression levels of HIF-1α and PPARGC1 and, most
increase in PPARGC1 and HIF-1α expression, RAP provoked
importantly, demonstrate that the compromised microcirculatory
profound changes in the ventricular expression of important
flow leads to ischemia-like conditions. The latter activate multiple
metabolic genes including hexokinase 2 (HK2), glycogen syn-
signaling pathways that are aimed at the improvement of oxygen
thase kinase 3β (GSK-3β), muscle isoform of glycogen phospho-
supply, angiogenesis, cell survival, and adaption of metabolism
rylase (PYGM), and acyl-coenzyme A dehydrogenase (ACADL;
Interestingly, these metabolic changes werenot affected by dronedarone, which suggests that other factors
than deprivation of oxygen and nutrients contribute to the overall
AT induces oxidative stress in the atrial and ventricular
change of ventricular gene expression during AF.
myocardium. In the ventricles, AF causes alterations in gene
It is fully established that even mild ischemia is associated
expression and activation of specific signal transduction pathways.
with compromised mitochondrial function and requires meta-
As a consequence, microcirculation is impaired, troponin can be
bolic adaption to maintain adequate ATP generation and cardiac
released, which is associated with causing clinical symptoms like
output Again, the activation of redox-
angina pectoris and dyspnea. It remains to be determined if these
sensitive transcription factors, namely HIF-1α and PPARGC1, is
alterations are also related to the increased rate of death in AF
responsible for these protective changes in the metabolism that
patients shown by several epidemiologic studies.
C. W. (2002). Elevated expression
NADPH oxidase. Arch. Biochem.
Abidov, A., Hachamovitch, R., Rozan-
of hexokinase II protects human
Biophys. 397, 342–324.
ski, A., Hayes, S. W., Santos, M. M.,
lung epithelial-like A549 cells against
Bayraktutan, U., Blayney, L., and
expression. J. Biol. Chem. 278,
Sciammarella, M. G., Cohen, I., Ger-
oxidative injury. Am. J. Physiol. Lung
Shah, A. M. (2000). Molecular
lach, J., Friedman, J. D., Germano,
Cell Mol. Physiol. 283, L573–L584.
characterization and localization
G., and Berman, D. S. (2004). Prog-
Amiri, F., Virdis, A., Neves, M. F.,
of the NAD(P)H oxidase com-
Guthrie, P. H., Carmical, S. G.,
nostic implications of atrial fibrilla-
Iglarz, M., Seidah, N. G., Touyz,
ponents gp91-phox and p22-phox
Chen, T. M., and Taegtmeyer, H.
tion in patients undergoing myocar-
R. M., Reudelhuber, T. L., and
in endothelial cells. Arterioscler.
(1996). Metabolic fate of glucose in
dial perfusion single-photon emis-
Schiffrin, E. L. (2004). Endothelium-
Thromb. Vasc. Biol. 20, 1903–1911.
reversible low-flow ischemia of the
sion computed tomography. J. Am.
restricted overexpression of human
Bendall, J. K., Cave, A. C., Heymes, C.,
isolated working rat heart. Am. J.
Coll. Cardiol. 44, 1062–1070.
endothelin-1 causes vascular remod-
Gall, N., and Shah, A. M. (2002). Piv-
Physiol. 270(Pt 2), H817–H826.
ACTIVE I Investigators, Yusuf, S.,
eling and endothelial dysfunction.
otal role of a gp91(phox)-containing
Bonello, S., Zahringer, C., BelAiba, R.
Healey, J. S., Pogue, J., Chrolavicius,
Circulation 110, 2233–2240.
NADPH oxidase in angiotensin II-
S., Djordjevic, T., Hess, J., Michiels,
S., Flather, M., Hart, R. G., Hohn-
Antoniades, C., Demosthenous, M.,
induced cardiac hypertrophy in
C., Kietzmann, T., and Görlach,
loser, S. H., Joyner, C. D., Pfeffer, M.
Reilly, S., Margaritis, M., Zhang, M.
mice. Circulation 105, 293–296.
A. (2007). Reactive oxygen species
A., and Connolly, S. J. (2011). Irbe-
H., Antonopoulos, A., Marinou, K.,
Berndt, C., Lillig, C. H., and Holmgren,
activate the HIF-1alpha promoter
sartan in patients with atrial fibrilla-
Nahar, K., Jayaram, R., Tousoulis,
A. (2007). Thiol-based mechanisms
via a functional NFkappaB site.
tion. N. Engl. J. Med. 364, 928–938.
D., Bakogiannis, C., Sayeed, R., Tri-
of the thioredoxin and glutaredoxin
Arterioscler. Thromb. Vasc. Biol. 27,
Afanas'ev, I. (2011). ROS and RNS
antafyllou, C., Koumallos, N., Psar-
systems: implications for diseases
signaling in heart disorders: could
ros, C., Miliou, A., Stefanadis, C.,
in the cardiovascular system. Am.
Brown, A. M., Sease, K. L., Robey, J.
antioxidant treatment be success-
Channon, K. M., and Casadei, B.
J. Physiol. Heart Circ. Physiol. 292,
L., Shofer, F. S., and Hollander, J.
ful? Oxid. Med. Cell. Longev. 2011,
(2012). Myocardial redox state pre-
E. (2007). The risk for acute coro-
293769. doi: 10.1155/2011/293769
dicts in-hospital clinical outcome
Besse, S., Tanguy, S., Riou, B., Boucher,
nary syndrome associated with atrial
Ago, T., Kuroda, J., Pain, J., Fu, C.,
after cardiac surgery effects of short-
F., Bulteau, A. L., Le Page, C., Swyn-
fibrillation among ED patients with
Li, H., and Sadoshima, J. (2010).
term pre-operative statin treatment.
ghedauw, B., and de Leiris, J. (2001).
chest pain syndromes. Am. J. Emerg.
Upregulation of NOX4 by hyper-
J. Am. Coll. Cardiol. 59, 60–70.
Coronary and aortic vasoreactivity
Med. 25, 523–528.
trophic stimuli promotes apopto-
Arany, Z., Foo, S. Y., Ma, Y., Ruas, J.
protection with endothelin receptor
Bruneau, B. G., Piazza, L. A., and de
sis and mitochondrial dysfunction
L., Bommi-Reddy, A., Girnun, G.,
antagonist, bosentan, after ischemia
Bold, A. J. (1997). BNP gene expres-
in cardiac myocytes. Circ. Res. 106,
Cooper, M., Laznik, D., Chinsom-
and hypoxia in aged rats. Eur. J.
sion is specifically modulated by
boon, J., Rangwala, S. M., Baek, K.
Pharmacol. 432, 167–175.
stretch and ET-1 in a new model
Ago, T., Liu, T., Zhai, P., Chen, W., Li,
H., Rosenzweig, A., and Spiegelman,
Bloom, D. A., and Jaiswal, A. K. (2003).
of isolated rat atria. Am. J. Physiol.
H., Molkentin, J. D., Vatner, S. F.,
B. M. (2008). HIF-independent reg-
Phosphorylation of Nrf2 at Ser40
273(Pt 2), H2678–H2686.
and Sadoshima, J. (2008). A redox-
ulation of VEGF and angiogen-
by protein kinase C in response to
Brunner, F., Leonhard, B., Kukovetz,
dependent pathway for regulating
esis by the transcriptional coac-
antioxidants leads to the release of
W. R., and Mayer, B. (1997).
class II HDACs and cardiac hyper-
tivator PGC-1alpha. Nature 451,
Nrf2 from INrf2, but is not required
trophy. Cell 133, 978–993.
for Nrf2 stabilization/accumulation
oxide and L-arginine release in
Ahmad, A., Ahmad, S., Schneider, B. K.,
Babior, B. M., Lambeth, J. D., and
in the nucleus and transcriptional
ischaemia/reperfusion injury of rat
Allen, C. B., Chang, L. Y., and White,
Nauseef, W. (2002). The neutrophil
activation of antioxidant response
heart. Cardiovasc. Res. 36, 60–66.
Goette et al.
Ventricular oxidative stress during AF
Bukowska, A., Hammwöhner, M., Six-
Lanas, F., Lewis, B. S., Merino, J. L.,
polycystic human ovaries. Am. J.
abnormalities in the ventricles. Eur.
dorf, A., Schild, L., Wiswedel, I.,
Morillo, C., Murin, J., Narasimhan,
Pathol. 162, 1881–1893.
Heart J. 30, 1411–1420.
Röhl, F. W., Wolke, C., Lendeckel,
C., Paolasso, E., Parkhomenko, A.,
Fineschi, M., Bravi, A., and Gori,
Goette, A., Bukowska, A., Lendeckel,
U., Aderkast, C., Bochmann, S.,
Peters, N. S., Sim, K. H., Stiles, M.
T. (2008). The "slow coronary
U., Erxleben, M., Hammwohner,
Chilukoti, R. K., Mostertz, J., Bram-
K., Tanomsup, S., Toivonen, L., Tom-
flow" phenomenon: evidence of
M., Strugala, D., Pfeiffenberger, J.,
lage, P., and Goette, A. (2012).
csányi, J., Torp-Pedersen, C., Tse, H.
preserved coronary flow reserve
Rohl, F. W., Huth, C., Ebert, M. P.,
Dronedarone prevents microcircu-
F., Vardas, P., Vinereanu, D., Xavier,
despite increased resting microvas-
Klein, H. U., and Rocken, C. (2008).
latory abnormalities in the left ven-
D., Zhu, J., Zhu, J. R., Baret-Cormel,
cular resistances. Int. J. Cardiol. 127,
Angiotensin II receptor blockade
tricle during atrial tachypacing. Br.
L., Weinling, E., Staiger, C., Yusuf,
reduces tachycardia-induced atrial
J. Pharmacol. 166, 964–980.
S., Chrolavicius, S., Afzal, R., Hohn-
Fontayne, A., Dang, P. M., Gougerot-
adhesion molecule expression. Cir-
Bukowska, A., Schild, L., Keilhoff, G.,
loser, S. H., and PALLAS Investiga-
Pocidalo, M. A., and El-Benna, J.
culation 117, 732–742.
Hirte, D., Neumann, M., Garde-
tors. (2011). Dronedarone in high-
(2002). Phosphorylation of p47phox
Goette, A., Staack, T., Rocken, C., Arndt,
mann, A., Neumann, K. H., Röhl,
risk permanent atrial fibrillation. N.
sites by PKC alpha, beta II, delta, and
M., Geller, J. C., Huth, C., Ansorge,
F. W., Huth, C., Goette, A., and
Engl. J. Med. 365, 2268–2276.
zeta: effect on binding to p22phox
S., Klein, H. U., and Lendeckel,
Lendeckel, U. (2008). Mitochondr-
Dammanahalli, K. J., and Sun, Z. (2008).
and on NADPH oxidase activation.
U. (2000). Increased expression of
ial dysfunction and redox signaling
Endothelins and NADPH oxidases
Biochemistry 41, 7743–7750.
extracellular signal-regulated kinase
in atrial tachyarrhythmia. Exp. Biol.
in the cardiovascular system. Clin.
Fuster, V., Ryden, L. E., Cannom, D. S.,
and angiotensin-converting enzyme
Med. (Maywood) 233, 558–574.
Exp. Pharmacol. Physiol. 35, 2–6.
Crijns, H. J., Curtis, A. B., Ellenbo-
in human atria during atrial fib-
Cadenas, E., Wefers, H., Müller, A.,
De Boer, R. A., Pinto, Y. M., and Van
gen, K. A., Halperin, J. L., Le Heuzey,
rillation. J. Am. Coll. Cardiol. 35,
Brigelius, R., and Sies, H. (1982).
Veldhuisen, D. J. (2003). The imbal-
J. Y., Kay, G. N., Lowe, J. E., Olsson,
Active oxygen metabolites and their
ance between oxygen demand and
S. B., Prystowsky, E. N., Tamargo, J.
Griendling, K. K., Sorescu, D., and
action in the hepatocyte. Studies on
supply as a potential mechanism in
L., Wann, S., Smith, S. C. Jr., Jacobs,
Ushio-Fukai, M. (2000). NAD(P)H
chemiluminescence responses and
the pathophysiology of heart failure:
A. K., Adams, C. D., Anderson, J. L.,
oxidase: role in cardiovascular biol-
alkane production. Agents Actions
the role of microvascular growth and
Antman, E. M., Halperin, J. L., Hunt,
ogy and disease. Circ. Res. 86,
Suppl. 11, 203–216.
abnormalities. Microcirculation 10,
S. A., Nishimura, R., Ornato, J. P.,
Camici, P. G., and Crea, F. (2007). Coro-
Page, R. L., Riegel, B., Priori, S. G.,
Hasdai, D., Kornowski, R., and Battler,
nary microvascular dysfunction. N.
Dikalova, A., Clempus, R., Lassègue,
Blanc, J. J., Budaj, A., Camm, A. J.,
A. (1994). Endothelin and myocar-
Engl. J. Med. 356, 830–840.
B., Cheng, G., McCoy, J., Dikalov,
Dean, V., Deckers, J. W., Despres, C.,
dial ischemia. Cardiovasc. Drugs
Cardin, S., Li, D., Thorin-Trescases,
S., San Martin, A., Lyle, A., Weber,
Dickstein, K., Lekakis, J., McGregor,
Ther. 8, 589–599.
N., Leung, T. K., Thorin, E., and
D. S., Weiss, D., Taylor, W. R.,
K., Metra, M., Morais, J., Osterspey,
Heusch, G. (2008). Heart rate in the
Nattel, S. (2003). Evolution of
Schmidt, H. H., Owens, G. K.,
A., Tamargo, J. L., and Zamorano,
pathophysiology of coronary blood
the atrial fibrillation substrate in
Lambeth, J. D., and Griendling, K.
J. L. (2006). ACC/AHA/ESC 2006
flow and myocardial ischaemia:
experimental congestive heart fail-
K. (2005). NOX1 overexpression
Guidelines for the Management of
benefit from selective bradycardic
ure: angiotensin-dependent and -
potentiates angiotensin II-induced
Patients with Atrial Fibrillation: a
independent pathways. Cardiovasc.
hypertension and vascular smooth
report of the American College of
Res. 60, 315–325.
muscle hypertrophy in transgenic
Cardiology/American Heart Associ-
Heusch, G., Baumgart, D., Cam-
Carnes, C. A., Chung, M. K., Nakayama,
mice. Circulation 112, 2668–2676.
ation Task Force on Practice Guide-
ici, P., Chilian, W., Gregorini, L.,
T., Nakayama, H., Baliga, R. S.,
Doerries, C., Grote, K., Hilfiker-Kleiner,
lines and the European Society of
Hess, O., Indolfi, C., and Rimoldi,
Piao, S., Kanderian, A., Pavia, S.,
D., Luchtefeld, M., Schaefer, A., Hol-
Cardiology Committee for Practice
O. (2000). Alpha-adrenergic coro-
Hamlin, R. L., McCarthy, P. M.,
land, S. M., Sorrentino, S., Manes,
Guidelines (Writing Committee to
nary vasoconstriction and myocar-
Bauer, J. A., and Van Wagoner,
C., Schieffer, B., Drexler, H., and
Revise the 2001 Guidelines for the
dial ischemia in humans. Circulation
D. R. (2001). Ascorbate attenuates
Landmesser, U. (2007). Critical role
Management of Patients With Atrial
101, 689–694.
atrial pacing-induced peroxynitrite
of the NAD(P)H oxidase subunit
Fibrillation): developed in collab-
H., Roder, F., Adami,
formation and electrical remodeling
p47phox for left ventricular remod-
oration with the European Heart
P., Voll, S., Kowallik, P., Haase,
and decreases the incidence of post-
eling/dysfunction and survival after
Rhythm Association and the Heart
A., Ertl, G., and Bauer, W. R.
operative atrial fibrillation. Circ. Res.
myocardial infarction. Circ. Res. 100,
Rhythm Society. Circulation 114,
(1997). Study of microcirculation by
89, E32–E38.
coloured microspheres and NMR-
Chen, H., Li, D., Sawamura, T., Inoue, K.,
Doughan, A. K., Harrison, D. G.,
Galatius-Jensen, S., Wroblewski, H.,
microscopy in isolated rat heart:
and Mehta, J. L. (2000). Upregula-
and Dikalov, S. I. (2008). Mole-
Emmeluth, C., Bie, P., Haunsø,
effect of ischaemia, endothelin-1
tion of LOX-1 expression in aorta of
cular mechanisms of angiotensin
S., and Kastrup, J. (1996). Plasma
and endothelin-1 antagonist BQ
hypercholesterolemic rabbits: mod-
II mediated mitochondrial dysfunc-
610. J. Mol. Cell. Cardiol. 29,
ulation by losartan. Biochem. Bio-
tion. Linking mitochondrial oxida-
failure: effect of the ACE inhibitor,
phys. Res. Commun. 276, 1100–1104.
tive damage and vascular endothe-
Hingtgen, S. D., Tian, X., Yang, J., Dun-
Chen, K., Kirber, M. T., Xiao, H., Yang,
lial dysfunction. Circ. Res. 102,
lay, S. M., Peek, A. S., Wu, Y., Sharma,
Y., and Keaney, J. F. Jr. (2008). Reg-
Ghezzi, P., Bonetto, V., and Fratelli,
R. V., Engelhardt, J. F., and Davisson,
ulation of ROS signal transduction
Engelhorn, T., Schwarz, M. A., Heusch,
M. (2005). Thiol-disulfide balance:
by NADPH oxidase 4 localization. J.
G., Doerfler, A., and Schulz, R.
from the concept of oxidative stress
NADPH oxidase and Akt activation
Cell Biol. 181, 1129–1139.
(2011). Reduction of cerebral infarct
to that of redox regulation. Antioxid.
play a key role in angiotensin
Connolly, S. J., Camm, A. J., Halperin, J.
size by dronedarone. Cardiovasc.
Redox Signal. 7, 964–972.
II-induced cardiomyocyte hyper-
L., Joyner, C., Alings, M., Amerena,
Drugs Ther. 25, 523–529.
Goette, A., Bukowska, A., Dobrev, D.,
J., Atar, D., Avezum, Á., Blomström,
Ferrara, N., Frantz, G., LeCouter, J.,
Pfeiffenberger, J., Morawietz, H.,
P., Borggrefe, M., Budaj, A., Chen,
Dillard-Telm, L., Pham, T., Drak-
Strugla, D., Wiswedel, I., Röhl, F.-W.,
Hohnloser, S. H., Crijns, H. J., van
S. A., Ching, C. K., Commerford, P.,
sharapu, A., Giordano, T., and
Wolke, C., Bergmann, S., Bramlage,
Eickels, M., Gaudin, C., Page, R. L.,
Dans, A., Davy, J. M., Delacrétaz, E.,
Peale, F. (2003). Differential expres-
P., Ravens, U., and Lendeckel, U.
Torp-Pedersen, C., Connolly, S. J.,
Di Pasquale, G., Diaz, R., Dorian,
sion of the angiogenic factor genes
(2009). Acute atrial tachyarrhyth-
and ATHENA Investigators. (2009).
P., Flaker, G., Golitsyn, S., Gonzalez-
vascular endothelial growth fac-
mia induces angiotensin II type
Effect of dronedarone on cardiovas-
Hermosillo, A., Granger, C. B., Hei-
tor (VEGF) and endocrine gland-
cular events in atrial fibrillation. N.
dbüchel, H., Kautzner, J., Kim, J. S.,
derived VEGF in normal and
Engl. J. Med. 360, 668–678.
Goette et al.
Ventricular oxidative stress during AF
Jaimes, E. A., Galceran, J. M., and Raij,
Karanikis, P., Pappa, E., Siogas, K.,
Martin, D., Galisteo, R., and Gutkind,
and Nattel, S. (2011). Mechanisms
L. (1998). Angiotensin II induces
and Goudevenos, J. A. (2005). Oral
J. S. (2009). CXCL8/IL8 stimu-
of atrial tachyarrhythmias associated
superoxide anion production by
vitamin C administration reduces
lates vascular endothelial growth
with coronary artery occlusion in
mesangial cells. Kidney Int. 54,
early recurrence rates after electri-
factor (VEGF) expression and the
a chronic canine model. Circulation
cal cardioversion of persistent atrial
autocrine activation of VEGFR2
123, 137–146.
Jones, D. P. (2006). Redefining oxida-
fibrillation and attenuates associated
in endothelial cells by activat-
tive stress. Antioxid. Redox Signal. 8,
inflammation. Int. J. Cardiol. 102,
ing NFkappaB through the CBM
Laubriet, A., and Rochette, L.
(Carma3/Bcl10/Malt1) complex. J.
(2003). Angiotensin II activates
Jones, D. P. (2008). Radical-free biology
Kunsch, C., and Medford, R. M. (1999).
Biol. Chem. 284, 6038–6042.
NADPH oxidase in isolated rat
of oxidative stress. Am. J. Physiol. Cell
Oxidative stress as a regulator of
Matsuno, K., Yamada, H., Iwata, K.,
Physiol. 295, C849–C868.
gene expression in the vasculature.
Jin, D., Katsuyama, M., Matsuki, M.,
reperfusion. Eur. J. Pharmacol. 462,
Kern, M. J., Lerman, A., Bech, J. W., De
Circ. Res. 85, 753–766.
Takai, S., Yamanishi, K., Miyazaki,
Bruyne, B., Eeckhout, E., Fearon, W.
Kuroda, J., Ago, T., Matsushima, S., Zhai,
M., Matsubara, H., and Yabe-
Qiu, X. B., Shao, Y. M., Miao, S., and
F., Higano, S. T., Lim, M. J., Meuwis-
P., Schneider, M. D., and Sadoshima,
Nishimura, C. (2005). NOX1 is
Wang, L. (2006). The diversity of
sen, M., Piek, J. J., Pijls, N. H., Siebes,
J. (2010). NADPH oxidase 4 (NOX4)
involved in angiotensin II-mediated
the DnaJ/Hsp40 family, the crucial
M., and Spaan, J. A. (2006). Physio-
is a major source of oxidative stress
hypertension: a study in NOX1-
partners for Hsp70 chaperones. Cell.
logical assessment of coronary artery
in the failing heart. Proc. Natl. Acad.
deficient mice. Circulation 112,
Mol. Life Sci. 63, 2560–2570.
disease in the cardiac catheterization
Sci. U.S.A. 107, 15565–15570.
Range, F. T., Schafers, M., Acil, T.,
laboratory: a scientific statement
Lawrence, D. M., Seth, P., Durham, L.,
Matsuzawa, A., and Ichijo, H. (2008).
Schafers, K. P., Kies, P., Paul, M., Her-
from the American Heart Associa-
Diaz, F., Boursiquot, R., Ransohoff,
Redox control of cell fate by MAP
mann, S., Brisse, B., Breithardt, G.,
tion Committee on Diagnostic and
R. M., and Major, E. O. (2006).
kinase: physiological roles of ASK1-
Schober, O., and Wichter, T. (2007).
Interventional Cardiac Catheteriza-
MAP kinase pathway in stress sig-
Impaired myocardial perfusion and
tion, Council on Clinical Cardiology.
tively upregulates CCL2/monocyte
naling. Biochim. Biophys. Acta 1780,
perfusion reserve associated with
Circulation 114, 1321–1341.
increased coronary resistance in per-
Kim, J. W., Tchernyshyov, I., Semenza,
Mayyas, F., Niebauer, M., Zurick,
sistent idiopathic atrial fibrillation.
G. L., and Dang, C. V. (2006). HIF-
progenitor cells. Glia 53, 81–91.
A., Barnard, J., Gillinov, A. M.,
Eur. Heart J. 28, 2223–2230.
1-mediated expression of pyruvate
Li, J. M., Gall, N. P., Grieve, D. J., Chen,
Chung, M. K., and Van Wag-
Reilly, S. N., Jayaram, R., Nahar, K.,
dehydrogenase kinase: a metabolic
M., and Shah, A. M. (2002). Activa-
oner, D. R. (2010). Association of
Antoniades, C., Verheule, S., Chan-
switch required for cellular adap-
tion of NADPH oxidase during pro-
left atrial endothelin-1 with atrial
non, K. M., Alp, N. J., Schotten,
tation to hypoxia. Cell Metab. 3,
gression of cardiac hypertrophy to
rhythm, size, and fibrosis in patients
U., and Casadei, B. (2011). Atrial
failure. Hypertension 40, 477–484.
with structural heart disease. Circ.
sources of reactive oxygen species
Kim, Y. M., Kattach, H., Ratnatunga,
Li, J. M., and Shah, A. M. (2003).
Arrhythm. Electrophysiol. 3, 369–379.
vary with the duration and substrate
C., Pillai, R., Channon, K. M., and
ROS generation by nonphagocytic
Mollnau, H., Wendt, M., Szöcs, K.,
of atrial fibrillation: implications for
Casadei, B. (2008). Association of
NADPH oxidase: potential relevance
Lassègue, B., Schulz, E., Oelze, M.,
the antiarrhythmic effect of statins.
atrial nicotinamide adenine dinu-
in diabetic nephropathy. J. Am. Soc.
Li, H., Bodenschatz, M., August, M.,
Circulation 124, 1107–1117.
cleotide phosphate oxidase activity
Nephrol. 14(Suppl. 3), S221–S226.
Kleschyov, A. L., Tsilimingas, N.,
Rinckel, L. A., Faris, S. L., Hitt, N. D.,
with the development of atrial fib-
Li, X., Zima, A. V., Sheikh, F., Blat-
Walter, U., Förstermann, U., Mein-
and Kleinberg, M. E. (1999). Rac1
rillation after cardiac surgery. J. Am.
ter, L. A., and Chen, J. (2005).
ertz, T., Griendling, K., and Münzel,
disrupts p67phox/p40phox binding:
Coll. Cardiol. 51, 68–74.
T. (2002). Effects of angiotensin II
a novel role for Rac in NADPH oxi-
Kinoshita, O., Yoshimi, H., Nagata, S.,
genic Ca2+ signaling is abolished
infusion on the expression and func-
dase activation. Biochem. Biophys.
Ishikura, F., Kimura, K., Yamabe,
in atrial myocytes of inositol-1,4,5-
tion of NAD(P)H oxidase and com-
Res. Commun. 263, 118–122.
T., Takagaki, K., Miyatake, K., and
ponents of nitric oxide/cGMP sig-
Rivard, L., Sinno, H., Shiroshita-
Omae, T. (1993). Rapid increase
2-deficient mice. Circ. Res. 96,
naling. Circ. Res. 90, E58–E65.
Takeshita, A., Schram, G., Leung,
in plasma endothelin concentrations
Murdoch, C. E., Zhang, M., Cave, A.
T. K., and Nattel, S. (2007).
during percutaneous balloon dilata-
Li, Y. L., Gao, L., Zucker, I. H., and
C., and Shah, A. M. (2006). NADPH
The pharmacological response of
tion of the mitral valve in patients
Schultz, H. D. (2007). NADPH
oxidase-dependent redox signalling
ischemia-related atrial fibrillation
with mitral stenosis. Br. Heart J. 69,
oxidase-derived superoxide anion
in cardiac hypertrophy, remodel-
in dogs: evidence for substrate-
mediates angiotensin II-enhanced
ling and failure. Cardiovasc. Res. 71,
specific efficacy. Cardiovasc. Res. 74,
Kiriakidis, S., Andreakos, E., Monaco,
carotid body chemoreceptor sensi-
C., Foxwell, B., Feldmann, M., and
tivity in heart failure rabbits. Cardio-
Nagase, T., Fukuchi, Y., Jo, C., Ter-
Rubens, C., Ewert, R., Halank, M.,
Paleolog, E. (2003). VEGF expres-
vasc. Res. 75, 546–554.
amoto, S., Uejima, Y., Ishida, K.,
Wensel, R., Orzechowski, H. D.,
sion in human macrophages is NF-
Lillig, C. H., and Holmgren, A. (2007).
Shimizu, T., and Orimo, H. (1990).
Schultheiss, H. P., and Hoeffken,
kappaB-dependent: studies using
Thioredoxin and related molecules –
Endothelin-1 stimulates arachido-
G. (2001). Big endothelin-1 and
adenoviruses expressing the endoge-
from biology to health and disease.
nate 15-lipoxygenase activity and
endothelin-1 plasma levels are cor-
nous NF-kappaB inhibitor Ikappa-
Antioxid. Redox Signal. 9, 25–47.
oxygen radical formation in the rat
related with the severity of primary
Balpha and a kinase-defective form
Lin, Y. K., Lai, M. S., Chen, Y. C., Cheng,
distal lung. Biochem. Biophys. Res.
pulmonary hypertension. Chest 120,
of the IkappaB kinase 2. J. Cell. Sci.
C. C., Huang, J. H., Chen, S. A.,
Commun. 168, 485–489.
116, 665–674.
Chen, Y. J., and Lin, C. I. (2012).
Neubauer, S., Zimmermann, S., Hirsch,
Satoh, M., Ogita, H., Takeshita, K.,
Kochiadakis, G. E., Skalidis, E. I., Kale-
Hypoxia and reoxygenation mod-
A., Pulzer, F., Tian, R., Bauer, W.,
Mukai, Y., Kwiatkowski, D. J., and
bubas, M. D., Igoumenidis, N. E.,
ulate the arrhythmogenic activity
Bauer, B., and Ertl, G. (1991).
Liao, J. K. (2006). Requirement of
Chrysostomakis, S. I., Kanoupakis,
of the pulmonary vein and atrium.
Effects of endothelin-1 in the iso-
Rac1 in the development of cardiac
E. M., Simantirakis, E. N., and Var-
Clin. Sci. 122, 121–132.
lated heart in ischemia/reperfusion
hypertrophy. Proc. Natl. Acad. Sci.
das, P. E. (2002). Effect of acute
Love, M. P., Ferro, C. J., Haynes, W.
and hypoxia/reoxygenation injury. J.
U.S.A. 103, 7432–7437.
atrial fibrillation on phasic coronary
G., Plumpton, C., Davenport, A. P.,
Mol. Cell. Cardiol. 23, 1397–1409.
Seagroves, T. N., Ryan, H. E., Lu, H.,
blood flow pattern and flow reserve
Webb, D. J., and McMurray, J. J.
Nishida, K., Qi, X. Y., Wakili, R., Com-
Wouters, B. G., Knapp, M., Thibault,
in humans. Eur. Heart J. 23, 734–741.
(2000). Endothelin receptor antag-
tois, P., Chartier, D., Harada, M.,
P., Laderoute, K., and Johnson, R. S.
Korantzopoulos, P., Kolettis, T. M.,
onism in patients with chronic heart
Iwasaki, Y. K., Romeo, P., Maguy, A.,
(2001). Transcription factor HIF-1 is
Kountouris, E., Dimitroula, V.,
failure. Cardiovasc. Res. 47, 166–172.
Dobrev, D., Michael, G., Talajic, M.,
a necessary mediator of the Pasteur
Goette et al.
Ventricular oxidative stress during AF
effect in mammalian cells. Mol. Cell.
Takahashi, N., Ishibashi, Y., Shimada,
Wyse, D. G., Waldo, A. L., DiMarco, J.
endothelin receptors in chronic
Biol. 21, 3436–3444.
T., Sakane, T., Ohata, S., Sug-
P., Domanski, M. J., Rosenberg, Y.,
Serrander, L., Cartier, L., Bedard, K.,
amori, T., Ohta, Y., Inoue, S.,
Schron, E. B., Kellen, J. C., Greene,
Banfi, B., Lardy, B., Plastre, O.,
Nakamura, K., Shimizu, H., Katoh,
H. L., Mickel, M. C., Dalquist, J. E.,
Sienkiewicz, A., Fórró, L., Schlegel,
H., and Murakami, Y. (2002).
and Corley, S. D. (2002). A com-
Conflict of Interest Statement: The
W., and Krause, K. H. (2007). NOX4
Impaired exercise-induced vasodi-
parison of rate control and rhythm
authors declare that the research was
activity is determined by mRNA lev-
latation in chronic atrial fibril-
control in patients with atrial fib-
conducted in the absence of any com-
els and reveals a unique pattern of
lation – role of endothelium-
rillation. N. Engl. J. Med. 347,
mercial or financial relationships that
ROS generation. Biochem. J. 406,
derived nitric oxide. Circ. J. 66,
could be construed as a potential con-
Yasunari, K., Maeda, K., Nakamura,
flict of interest.
Shohet, R. V., and Garcia, J. A. (2007).
Van Gelder, I. C., Hagens, V. E.,
M., and Yoshikawa, J. (2002).
Keeping the engine primed: HIF
Bosker, H. A., Kingma, J. H., Kamp,
Received: 22 January 2012; accepted: 11
factors as key regulators of car-
O., Kingma, T., Said, S. A., Dar-
II – mediated migration of human
June 2012; published online: 05 July 2012.
diac metabolism and angiogenesis
manata, J. I., Timmermans, A. J.,
Citation: Goette A, Bukowska A, Lillig
during ischemia. J. Mol. Med. 85,
Tijssen, J. G., and Crijns, H. J.
CH and Lendeckel U (2012) Oxidative
(2002). A comparison of rate con-
stress. Hypertension 39(2 Pt 2),
stress and microcirculatory flow abnor-
Sinno, H., Derakhchan, K., Libersan, D.,
trol and rhythm control in patients
malities in the ventricles during atrial
Merhi, Y., Leung, T. K., and Nattel,
with recurrent persistent atrial fib-
Zhang, Y., Griendling, K. K., Dikalova,
fibrillation. Front. Physio. 3:236. doi:
S. (2003). Atrial ischemia promotes
rillation. N. Engl. J. Med. 347,
A., Owens, G. K., and Taylor, W.
atrial fibrillation in dogs. Circulation
R. (2005). Vascular hypertrophy in
This article was submitted to Frontiers in
107, 1930–1936.
White, M., Rouleau, J. L., Hall, C.,
angiotensin II-induced hyperten-
Cardiac Electrophysiology, a specialty of
Skyschally, A., and Heusch, G. (2011).
Arnold, M., Harel, F., Sirois, P.,
sion is mediated by vascular smooth
Frontiers in Physiology.
Reduction of myocardial infarct
Greaves, S., Solomon, S., Ajani, U.,
muscle cell-derived H2O2. Hyper-
Copyright 2012 Goette, Bukowska, Lil-
size by dronedarone in pigs –
Glynn, R., Hennekens, C., and Pfef-
tension 46, 732–737.
lig and Lendeckel. This is an open-access
a pleiotropic action? Cardiovasc.
fer, M. (2001). Changes in vasocon-
Zolk, O., Quattek, J., Sitzler, G.,
article distributed under the terms of
Drugs Ther. 25, 197–201.
strictive hormones, natriuretic pep-
Schrader, T., Nickenig, G., Schn-
Sugden, P. H. (2003). An overview of
tides, and left ventricular remod-
abel, P., Shimada, K., Takahashi,
which permits non-
endothelin signaling in the cardiac
eling soon after anterior myocar-
commercial use, distribution, and repro-
myocyte. J. Mol. Cell. Cardiol. 35,
dial infarction. Am. Heart J. 142,
duction in other forums, provided the
endothelin-converting enzyme, and
original authors and source are credited.
Source: http://lillig.eu/data/fphys3(236)2012.pdf
A. Elmanama et al., J. Al-Aqsa Unv., 10 (S.E.) 2006 Contribution of Hospital Wastewater to the Spread of Antibiotic Resistance in Comparison to Non-Health Institution Dr. Abdelraouf A. Elmanama * Dr. Abboud Y. ElKichaoui ** Miss. Mai Mohsin ∗∗* ﺭﻴﻭـﻁﺘ ﻩﺎـﺠﺘﺍ ﻲﻓ ﺭﺍﺩﺤﻨﻼﻟ ﺔﻓﺎﻀ ﻹﺎﺒ ﺔﻴﻭﻴﺤﻟﺍ ﺕﺍﺩﺎﻀﻤﻠﻟ ﺔﻤﻭﺎﻘﻤﻟﺍ ﺔﻠﻜﺸﻤﻟ ﺔﻴﻤﺎﻌﻟﺍ ﺓﺩﺎﻴﺯﻟﺍ ﺎﻴﺭﻴﺘﻜﺒﻟﺍ ﺔﻤﻭﺎﻘﻤ ﺭﻭﺼ ﺔﺴﺍﺭﺩﻟ ﺙﺤﺒﻟﺍ ﺍﺫﻫ ﻡﻤﺼ .
Prof. Dr. Christian Schubel Sommersemester 2008 Europäisches Wettbewerbsrecht § 1 Das Kartellverbot – Tatbestand und Rechtsfolgen I. Sachlicher und örtlicher Anwendungsbereich das europäische Kartellrecht gilt umfassend es gibt also keine Bereichsausnahmen, insb. unterstehen auch Banken, Versicherungen und Versorgungsunternehmen dem europäischen Kartellrecht



