Viagra gibt es mittlerweile nicht nur als Original, sondern auch in Form von Generika. Diese enthalten denselben Wirkstoff Sildenafil. Patienten suchen deshalb nach viagra generika schweiz, um ein günstigeres Präparat zu finden. Unterschiede bestehen oft nur in Verpackung und Preis.
Transferable vancomycin resistance in a community-associated mrsa lineage
Transferable Vancomycin Resistance
in a Community-Associated MRSA Lineage
Flávia Rossi, M.D., Ph.D., Lorena Diaz, Ph.D., Aye Wollam, B.Sc.,
Diana Panesso, Ph.D., Yanjiao Zhou, Ph.D., Sandra Rincon, M.Sc.,
Apurva Narechania, M.A., Galen Xing, Thais S.R. Di Gioia, M.D., André Doi, M.D.,
Truc T. Tran, Pharm.D., Jinnethe Reyes, M.Sc., Jose M. Munita, M.D.,
Lina P. Carvajal, B.Sc., Alejandra Hernandez-Roldan, M.Sc., Denise Brandão, M.D.,
Inneke Marie van der Heijden, Ph.D., Barbara E. Murray, M.D.,
Paul J. Planet, M.D., Ph.D., George M. Weinstock, Ph.D.,
and Cesar A. Arias, M.D., Ph.D.
From the Department of Pathology, Divi-
We report the case of a patient from Brazil with a bloodstream infection caused by
sion of Microbiology of the Central Labo-
a strain of methicillin-resistant
Staphylococcus aureus (MRSA) that was susceptible to
ratory (LIM 03) and Department of In-fectious Disease (LIM 54), Hospital das vancomycin (designated BR-VSSA) but that acquired the
vanA gene cluster during Clínicas da Faculdade de Medicina da antibiotic therapy and became resistant to vancomycin (designated BR-VRSA). Both
Universidade de São Paulo, São Paulo strains belong to the sequence type (ST) 8 community-associated genetic lineage
(F.R., T.S.R.D.G., A.D., D.B., I.M.H.); Mo-lecular Genetics and Antimicrobial Resis-
that carries the staphylococcal chromosomal cassette
mec (SCC
mec) type IVa and the
tance Unit, Universidad El Bosque, Bogota
S. aureus protein A gene (
spa) type t292 and are phylogenetically related to MRSA
(L.D., D.P., S.R., J.R., L.P.C., C.A.A.), and lineage USA300. A conjugative plasmid of 55,706 bp (pBRZ01) carrying the
vanA
Centro Internacional de Entrena miento e Investigaciones Medicas (CIDEIM), Cali cluster was identified and readily transferred to other staphylococci. The pBRZ01 (A.H.-R.) — both in Colombia; Clinica plasmid harbors DNA sequences that are typical of the plasmid-associated replica-
Alemana de Santiago, Universidad del tion genes
rep24 or
rep21 described in community-associated MRSA strains from
Desarrollo, Santiago, Chile (J.M.M.); the Department of Internal Medicine, Division Australia (pWBG745). The presence and dissemination of community-associated of Infectious Diseases (L.D., D.P., T.T.T., MRSA containing
vanA could become a serious public health concern.
J.R., J.M.M., A.H.-R., B.E.M, C.A.A.), and the Department of Microbiology and Mo-lecular Genetics (B.E.M., C.A.A.), Univer-sity of Texas Medical School at Houston, and the University of Houston College of Since the description in 2002 of the
vanA gene cluster in MRSA as
the mechanism of high-level vancomycin resistance, 13 isolates have been re-
Pharmacy (T.T.T.) — both in Houston; the Genome Institute, Washington University
ported in the United States,1,2 with others reported in India3 and Iran.4
Entero
at St. Louis, St. Louis (A.W., Y.Z., G.M.W.);
coccus faecalis and
E. faecium have been implicated as the donors of the vancomycin
and Sackler Institute for Comparative Ge-
resistance genes.5,6 The VRSA isolates were recovered from patients with soft-tissue
nomics, American Museum of Natural His-tory (A.N., P.J.P.), and Division of Pediatric or skin infections, and multilocus sequence typing has shown that 12 U.S. VRSA iso-Infectious Diseases, Columbia University lates belong to clonal complex 5 (including ST5, ST85, ST231, and ST371)1 and that
(G.X., P.J.P.) — both in New York. Ad-
the 13th isolate belongs to clonal complex 30. Clonal complex 5 is the most widely
dress reprint requests to Dr. Arias at the University of Texas Medical School, 6431 disseminated hospital-associated MRSA clonal complex in the United States, with pat-Fannin St., Rm. 2.112 MSB, Houston, TX terns designated as USA100 and USA800 on pulsed-field gel electrophoresis (PFGE).1
77030, or at
[email protected].
Community-associated MRSA emerged during the 1990s and rapidly dissemi-
This article was updated on April 17, 2014, nated across the United States, most often causing severe skin and soft-tissue in-at NEJM.org.
fections but on occasion causing life-threatening infections such as necrotizing
pneumonia.7 Early variants of this community-associated MRSA were initially
N Engl J Med 2014;370:1524-31.
DOI: 10.1056/NEJMoa1303359
traced to PFGE pattern USA400, but these variants were rapidly replaced by isolates
Copyright 2014 Massachusetts Medical Society.
within the genetic lineage designated USA300-ST8 (clonal complex 8). A variant
of USA300-ST8 emerged in South America in 2005 (designated USA300 Latin
n engl j med 370;16 nejm.org april
The New England Journal of Medicine
Downloaded from nejm.org at FIOCRUZ on April 24, 2014. For personal use only. No other uses without permission.
Copyright 2014 Massachusetts Medical Society. All rights reserved.
Transferable Vancomycin Resistance in MRSA Lineage
American Variant [USA300-LV])8 and has spread recommendation of contact isolation. A sample
rapidly, replacing the previously predominant from a rectal swab obtained 2 weeks after the
clonal complex 5 hos pital-associated clone (known initial positive blood culture indicated coloniza-
as the Chilean–Cordobes clone).9,10 Here we re- tion with vancomycin-resistant
E. faecalis (VREF).
port on the acquisition of the
vanA gene cluster The fever diminished, and daptomycin was con-
by an invasive bloodstream isolate of community- tinued for an additional 4 weeks. However, while
associated MRSA lineage in a patient at a Brazilian the patient was receiving daptomycin, oral muco-
sal lesions developed and his respiratory status
and fever worsened. Blood cultures yielded
Steno
trophomonas maltophilia; treatment with levofloxa-
cin was started and intravenous catheters were
The patient was a 35-year-old man from São Paulo removed. Respiratory secretions yielded extended-
with mycosis fungoides, cocaine addiction, and spectrum, beta-lactamase–producing
Kleb siel a pneu
diabetes mellitus. He was initially admitted to a
moniae and carbapenem-resistant
Acinetobacter bau
psychiatric hospital in November 2011 for depres-
mannii. Polymixin B sulfate was added to the
sion and suicidal ideation. Leg cellulitis developed treatment regimen. Multisystem organ failure
and was treated with cephalexin and topical gen- ensued, and blood cultures grew
Candida albi
tamicin. The patient was discharged with a pre-
cans. Be cause of the patient's worsening status,
scription for clindamycin but was readmitted in care was withdrawn in November 2012. The
June 2012 because of worsening psychiatric symp- patient died while receiving meropenem, lin ez-
toms and recurrent infections of the skin and soft olid, polymyxin B sulfate, and amphotericin B.
tissues. Vancomycin and cefepime were adminis- (A summary of the patient's course is presented
tered for 8 days; blood cultures obtained during in Fig. 1.)
this period were sterile. The patient remained in
the hospital to receive chemotherapy with gem-
citabine, vinorelbine, and dexamethasone for my-
cosis fungoides (skin cancer). In July 2012, while
Bacterial Identification, Susceptibility
the patient remained in the hospital, fever recurred,
Testing, and Molecular Typing
and treatment with vancomycin and piperacillin–
S. aureus and VREF isolates recovered from the
tazobactam was started. On July 16 and 24, blood patient's blood samples and rectal swabs, respec-
cultures yielded MRSA isolates that were suscep- tively, were identified by means of a polymerase-
tible to vancomycin, linezolid, and clindamycin chain-reaction (PCR) assay11,12 and 16S ribosomal
(see Table S1 in the Supplementary Appendix, avail- RNA sequencing.13 Susceptibility was determined
able with the full text of this article at NEJM.org). with the use of agar dilution and broth microdilu-
Vancomycin was continued for 13 days (highest tion.14 To assess the genetic background of the
vancomycin trough level, 8.7 μg per milliliter), isolates, PFGE was performed on two
S. aureus iso-
followed by treatment with teicoplanin for an ad- lates recovered from the patient's blood (BR-VSSA
ditional 14 days.
and BR-VRSA) (Fig. 1); on representative MRSA
A transthoracic echocardiogram showed no evi- isolates circulating in South American hospitals,
dence of infective endocarditis. On August 15, including the Latin American variant (USA300-
the day after teicoplanin was discontinued, fever LV) and Chilean–Cordobes clones; and on refer-
recurred. Blood cultures were positive for two ence strains from the following pandemic clones:
MRSA isolates, one of which was resistant to USA300 (NRS482),9 the Brazilian clone (F338),
both vancomycin (minimal inhibitory concen- the Iberian clone (NRS385-USA500), the Pediat-
tration, >32 μg per milliliter) and teicoplanin ric clone (NRS387-USA800), and the New York–
(minimal inhibitory concentration, 32 μg per Japan clone (NRS382-USA100). Typing of BR-VRSA
milliliter); the isolate was also resistant to eryth- was performed with the use of multilocus se-
romycin, clin da mycin, ciprofloxacin, gentamicin, quence typing, SCC
mec, and
spa. PCR assays were
and trimethoprim–sulfamethoxazole (Table S1 in used to detect the genes encoding Panton–
the Supplementary Appendix).
Valentine leukocidin (PVL) and the arginine cata-
Treatment with daptomycin was started, and bolic mobile element, which are often found in
the patient was placed in a single room, with a MRSA USA300.9
n engl j med 370;16 nejm.org april
The New England Journal of Medicine
Downloaded from nejm.org at FIOCRUZ on April 24, 2014. For personal use only. No other uses without permission.
Copyright 2014 Massachusetts Medical Society. All rights reserved.
Plasmid Characterization and Bacterial
performed to classify plasmid replicons. The
Biosafety Committee at the University of Texas–
We assessed the transferability of resistance to Houston approved these experiments.
vancomycin by performing filter matings,15 us-
ing BR-VRSA and the VREF isolate from the pa-
Whole-Genome Sequencing and Phylogenetic
tient as donors and BR-VSSA,
S. aureus RN4220,
Analysis
S. aureus COL, and
E. faecalis OG1RF as recipients BR-VSSA, BR-VRSA, and VREF isolates were evalu-
(Table S2 in the Supplementary Appendix). A donor- ated by means of whole-genome sequencing with
to-recipient ratio of 1:10 was used for mating ex- the use of the Illumina platform. Annotation and
periments, as described previously.15 Selection was assembly were performed as previously described19;
performed on brain–heart infusion (BHI) agar details are available at the National Center for
supplemented with vancomycin and fusidic acid at Bio technology Information (GenBank Bioproject
concentrations of 32 μg per milliliter and 25 μg numbers 205852 [for BR-VSSA], PRJNA183704
per milliliter, respectively. To determine whether [for BR-VRSA], and 205838 [for VREF]). Details of
vanA was present on a plasmid, S1 nuclease di- genomic comparisons (including the identifica-
gestion of total DNA, coupled with PFGE and hy- tion of single-nucleotide polymorphisms [SNPs])
bridization with a
vanA probe, was performed.16,17 and phylogenetic analysis are described in the
PCR assays targeting pSK41-like6 and Inc18-like Supplementary Appendix.
plasmids6 and the
rep plasmid families18 were
A Novel Transferable vanA Plasmid in BR-VRSA
Initial molecular characterization showed that
BR-VRSA displayed a PFGE pattern that was in-
distinguishable from that of BR-VSSA (Fig. 2A), sug-
gesting that BR-VRSA was selected in vivo during
the administration of glycopeptides and other anti-
microbial drugs. Moreover, S1 nuclease digestion
and hybridization showed that BR-VRSA contained
a plasmid of approximately 55 kb (not present in
BR-VSSA) that carried
vanA and
aac(6')–aph(2''
),
which encode enzymes that confer resistance to
vancomycin and gentamicin, respectively (Fig. 2B).
Growth rates were similar for both organisms
(Fig. S1 in the Supplementary Appendix), suggest-
ing that acquisition of the
vanA plasmid did not
Nov. 2011–
Feb. 2012
affect the in vitro fitness of the strain.
Filter-mating assays showed that the rate of
transfer of vancomycin resistance from BR-VRSA
Figure 1. The Patient's Clinical Course before and after Isolation
of the Vancomycin-Resistant Staphylococcus aureus.
(donor) to BR-VSSA (recipient) was 2.6×10−4 trans-
The antibiotics used are depicted in colored rectangles, with beta-lactams
conjugants per donor (Table S2 in the Supple-
in blue (cephalexin, cefepime, and piperacillin–tazobactam), clindamycin in
men tary Appendix). Rates of transfer to other
green, glycopeptides in pink (vancomycin and teicoplanin), and daptomycin
staphylococci (
S. aureus RN4220-RF and COL)
in yellow; the number in each rectangle corresponds to the number of days
were 1.65×10−4 and 6×10−5, respectively (Table S2
of treatment with the antibiotic. The drugs are shown in the order in which
in the Supplementary Appendix). Experiments
they were added to therapy. Dosages were as follows: cephalexin, 500 mg every 6 hours, given orally; clindamycin, 450 mg every 8 hours, given orally;
with S1 nuclease indicated that the plasmid con-
cefepime, 1 g every 8 hours, given intravenously; piperacillin–tazobactam,
taining
vanA (designated pBRZ01; 55,706 bp)
3.375 g every 6 hours, given intravenously; vancomycin, 1 g every 12 hours,
was the only large plasmid that was readily
given intravenously; teicoplanin, 400 mg daily, given intravenously; and dapto-
transferred to
S. aureus during the mating experi-
mycin, 6 mg per kilogram of total body weight daily, given intravenously. The
ment (Fig. 2B). We were unable to transfer vanco-
final days of hospitalization are not included. SSTI denotes skin and soft-tissue infection, BR-VSSA vancomycin-susceptible
S. aureus, BR-VRSA vancomycin-
mycin resistance to a laboratory strain of
E. faeca
resistant
S. aureus, and VREF vancomycin-resistant
Enterococcus faecalis.
lis (strain OG1RF), which suggests that the
plasmid was not able to replicate in enterococci.
n engl j med 370;16 nejm.org april
The New England Journal of Medicine
Downloaded from nejm.org at FIOCRUZ on April 24, 2014. For personal use only. No other uses without permission.
Copyright 2014 Massachusetts Medical Society. All rights reserved.

Transferable Vancomycin Resistance in MRSA Lineage
The transfer of vancomycin resistance from inverted repeat (Fig. S2 in the Supplementary Ap-
VREF recovered from the patient's rectal swab pendix), was identified, with the insertion of two
was also unsuccessful.
open reading frames encoding an enterococcal
Analysis of the sequence of pBRZ01 (Fig. 2C) resolvase and transposase (from the Tn3 family),
indicated that a Tn1546-like element underwent respectively (Fig. S2 in the Supplementary Ap pen-
important DNA rearrangements (Fig. S2 in the dix). The sequence of the vanA cluster (including
Supplementary Appendix). An insertion sequence 614 bp upstream and 222 bp downstream) har-
(IS1216) was found at the 5' end of the Tn1546 bored by the VREF infecting the patient was
variant, with a deletion of 3397 bp eliminating the identical to the sequence in BR-VRSA and was
left inverted repeat, the gene encoding integrase also identical to the sequences from plasmids
(open reading frame 1), and part of the gene en- pWZ7140 and pWZ909, previously identified in
coding resolvase (open reading frame 2) (Fig. S2 E. faecalis.20 These findings support the view
in the Supplementary Appendix). Downstream of that the vanA cluster has an enterococcal origin.
vanZ, a deletion of 96 bp, including the right Genomic analysis (confirmed by means of PCR
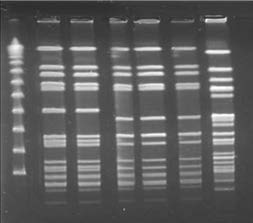
A Smal Digestion PFGE
B S1 Nuclease Digestion and Hybridization with VanA
C BR-VRSA Plasmid pBRZ01
Vancomycin resistance conferred by the vanA cluster
Aminoglycoside resistance conferred by acc(6')–aph(2'')
DNA replication, cell division, conjugation machinery
Transposases and resolvases
Hypothetical proteins
Figure 2. Results of Pulsed-Field Gel Electrophoresis (PFGE), S1 Nuclease Digestion of Total DNA, Hybridization,
and Plasmid Annotation.
Panel A shows the results of SmaI digestion of total DNA followed by PFGE. Lane M shows the lambda ladder (with
the molecular sizes in kilobases shown at left); lane 1, BR-VSSA; lane 2, BR-VRSA (donor); lane 3, transconjugant
number 1 Staphylococcus aureus RN4220-RF; lane 4, transconjugant number 2 S. aureus RN4220-RF; lane 5, S. aureus
RN4220-RF (recipient); and lane 6, vancomycin-resistant Enterococcus faecalis (vanA). Panel B shows the results of S1
digestion of the total DNA of staphylococcal strains and vancomycin-resistant E. faecalis followed by PFGE (at left) and
hybridization with a VanA probe (at right). In the results at left, lane 1 shows BR-VSSA; lane 2, BR-VRSA (donor);
lane 3, transconjugant number 1 S. aureus RN4220-RF; lane 4, transconjugant number 2 S. aureus RN4220-RF; lane 5,
S. aureus RN4220RF (recipient); lane 6, vancomycin-resistant E. faecalis; and lane M the lambda ladder. In the results at
right, the white and red arrows highlight the positive signal for vanA hybridization in staphylococcal and enterococcal
strains, respectively. Panel C shows a schematic representation of the annotation of the BR-VRSA plasmid pBRZ01.
n engl j med 370;16 nejm.org april
The New England Journal of Medicine
Downloaded from nejm.org at FIOCRUZ on April 24, 2014. For personal use only. No other uses without permission.
Copyright 2014 Massachusetts Medical Society. All rights reserved.
assay) of VREF indicated that the repR gene score of q>20) differentiating the two strains,
(typical of Inc18 type plasmids previously associ- whereas a comparison of each of these strains
ated with transfer of the vanA cluster into clonal with the USA300 strain TCH1516 showed a dif-
complex 5 S. aureus) and the traA gene (required ference of 1637 SNPs (965 core SNPs) for BR-VSSA
for plasmid transfer)6 were absent, which supports and 1757 SNPs (1005 core SNPs) for BR-VRSA,
the absence of transferability of the enterococcal suggesting that BR-VSSA is more similar to the
plasmid and further indicating that VREF was progenitor of BR-VRSA. (The SNP score is a mea-
not the direct donor of vanA genes to BR-VRSA. sure of the number of reads from a sequencing
Comparison of pBRZ01 with pWBG745 that agree with regard to a particular SNP.)
(38,204 bp), a plasmid identified in community-
associated MRSA isolates from remote areas of
Australia (clonal complex 5), showed 99% iden-
tity across 21,164 nucleotides.21 In addition, a The acquisition of high-level vancomycin resis-
sequence analysis indicated that the plasmid tance by S. aureus has been deemed a major clini-
containing vanA is not related to the pSK41-like cal and epidemiologic threat. However, since the
staphylococcal or Inc18-like enterococcal plasmids first characterization of a strain of VRSA in the
previously associated with vanA-carrying plasmids United States,5,23 only a few other isolates have
from VRSA isolates identified in the United States been reported. All the VRSA isolates from the
and belonging to clonal complex 5.1,6 Instead, the United States described to date have been isolated
vanA plasmid harbors sequences that are typical from infections of the skin and soft-tissue or
of the recently designated rep24 family18 or rep21 from colonizers of the skin. This case documents
family22 found in pWBG745. (Other genetic de- a bloodstream infection caused by a VRSA strain
terminants conferring antibiotic resistance that in a patient with a serious skin condition that
are present in the BR-VRSA genome are shown probably predisposed the patient to bacterial
in Table S3 in the Supplementary Appendix.)
colonization. The patient was treated repeatedly
with beta-lactams and glycopeptides, and our
ST8 Community-Associated Genetic
analyses suggest that the MRSA isolate had also
Background in BR-VRSA
acquired the vanA plasmid during therapy. The
PFGE indicated that BR-VRSA is genetically re- genetic characterization and genomic analyses
lated to MRSA USA300 (Fig. 3A), and multilocus also suggest that the vanA gene cluster present in
sequence typing showed that BR-VRSA belongs BR-VRSA may have originated from an entero-
to ST8 (as do USA300 and USA300-LV), harbors coccal donor. The DNA sequence of the vanA
SCCmec type IVa, and is spa type t292. Unlike gene cluster (and partial flanking sequences) of
USA300 and USA300-LV, BR-VRSA lacked genes BR-VRSA is identical to the sequence present in
encoding PVL. Neither the arcA gene nor the en- the genome of VREF that was recovered from a
tire arginine catabolic mobile element was swab of the patient's rectum, suggesting that
present in BR-VRSA, as has also been noted for VREF may have been the donor. However, the
MRSA strains belonging to the USA300-LV lin- presence of a different plasmid in this isolate and
eage. BR-VRSA and BR-VSSA harbor an intact the absence of transferability of the vanA cluster
bsa operon (for bacteriocin production) that is from VREF to staphylococci or enterococci suggest
characteristic of community-associated MRSA; that acquisition occurred through different inter-
VRSA strains from clonal complex 5 lack this mediaries. Indeed, the Tn1546-like element in
operon.1 Whole-genome, SNP-based phylogenetic BR-VRSA was altered, indicating that po tentially
analysis (performed with either the whole ge- important genetic rearrangements of Tn154624
nome [Fig. 3B] or the core genome [Fig. S3 in the had occurred. The presence of the insertion se-
Supplementary Appendix]) showed that there is a quences flanking the vanA gene cluster (Fig. S2 in
close relationship between BR-VSSA and BR-VRSA the Supplementary Appendix) may also provide the
and between each of these variants and other truncated Tn1546-derived element with mobility.
ST8 genomes, including USA300 and USA300-LV.
The results of our genomic and phylogenetic
Genomic comparisons of BR-VSSA and BR-VRSA analyses (with more SNP changes detected on
revealed only 288 SNPs (134 in the core genome) the branches leading to BR-VRSA than on those
with a score indicating high quality (defined as a leading to BR-VSSA) suggest that BR-VRSA is
n engl j med 370;16 nejm.org april
The New England Journal of Medicine
Downloaded from nejm.org at FIOCRUZ on April 24, 2014. For personal use only. No other uses without permission.
Copyright 2014 Massachusetts Medical Society. All rights reserved.

Transferable Vancomycin Resistance in MRSA Lineage
derived from BR-VSSA or a strain closely resem- Table S1 in the Supplementary Appendix) were
bling BR-VSSA. However, since BR-VSSA did not gentamicin-resistant and vancomycin-susceptible,
contain pBRZ01, the origin of this plasmid is and it is tempting to speculate that they may
unknown. The two MRSA isolates recovered (but have been carriers of a version of pBRZ01 that
then discarded) from the patient's blood before did not contain the vanA gene cluster.
the isolation of BR-VSSA and BR-VRSA (Fig. 1, and
Apart from the type of infection, several factors
A PFGE and Dendrogram of S. aureus Strains
USA300 (Nebraska)
Percentage of Similarity
B Phylogenetic Tree
0.03 substitutions
Figure 3. Phylogenetic Comparisons of BR-VRSA and BR-VSSA with Representatives of Other Methicillin-Resistant
Staphylococcus aureus (MRSA) Clones.
Panel A shows a dendrogram of the pandemic MRSA clones, BR-VRSA and BR-VSSA, generated with PFGE and the
use of GelCompar II software, version 6.5 (Applied Maths). Patterns were clustered by means of UPGMA (unweighted
pair group method with arithmetic mean), with the use of the Dice similarity coefficient and an optimization of 0.50%
and a tolerance of 1.0%. PFGE types, or clusters, were identified on the basis of a similarity of 75% or higher (indicated by
the vertical red line). ST denotes sequence type. The phylogenetic tree shown in Panel B is based on whole-genome
single-nucleotide polymorphisms (SNPs) and was generated with the use of the maximum-likelihood optimality
criterion. Branch lengths are proportional to the number of evolutionary changes (substitutions per site). All nodes
have 100% bootstrap support. Sequence types (STs) and clonal complexes (CCs) of the S. aureus strains are indicated.
n engl j med 370;16 nejm.org april
The New England Journal of Medicine
Downloaded from nejm.org at FIOCRUZ on April 24, 2014. For personal use only. No other uses without permission.
Copyright 2014 Massachusetts Medical Society. All rights reserved.
that may have important public health implications USA300 genetic lineage and capable of producing
distinguish this case from previously reported severe, invasive bloodstream infections. This obser-
cases of VRSA infection. First, this case is an ex- vation is supported by our finding that BR-VRSA,
ample of the acquisition of a vanA gene cluster that unlike previously reported VRSA strains, harbors
occurred independently of the hospital-associated an intact bsa operon,1 a genetic marker of com-
MRSA clonal complex 5 lineage. The molecular and munity-associated MRSA strains.26 In addition,
genomic data indicate that the genetic background characterization and sequencing of pBRZ01 indi-
of the VRSA strain is closely related to community- cate that rep24 or rep21 family plasmids recently
associated MRSA strains dis semi nat ed in several described in community-associated MRSA (belong-
parts of the world (USA300 ST8 harboring SCCmec ing to clon al complex 5)21 in Australia have been
type IVa). An ST8 community-associated variant of acquired by the MRSA ST8 genetic lineage and are
the MRSA USA300 clone (USA300-LV), which has capable of capturing the vanA cluster on a trans-
been reported in the northern part of South ferable plasmid. Finally, because USA300-like
America,8,9,25 has almost replaced the common strains spread rapidly and replace other MRSA
hospital-associated clone (ST5 Chilean–Cordobes lineages with such efficiency, our findings raise
clone).9,10,25 However, the genetic background of the possibility that vancomycin resistance may be
BR-VRSA differs from that of USA300-LV and from disseminated to MRSA clonal complex 8 isolates
that of the prototypical USA300 in that it lacks the in both the hospital and the community.
genes encoding PVL, a genetic marker of commu-
Supported by grants from the National Institute of Allergy and
nity-associated MRSA strains. This difference sug- Infectious Diseases (R01 AI093749, to Dr. Arias) and the National
gests that BR-VRSA is a novel variant of ST8 Institutes of Health (1U54 HG004968, to Dr. Weinstock).
Disclosure forms provided by the authors are available with
community-associated MRSA, closely related to the the full text of this article at NEJM.org.
1. Kos VN, Desjardins CA, Griggs A, et 8. Arias CA, Rincon S, Chowdhury S, 15. Tomita H, Pierson C, Lim SK, Clewell
al. Comparative genomics of vancomycin- et al. MRSA USA300 clone and VREF — DB, Ike Y. Possible connection between
resistant Staphylococcus aureus strains and a U.S.–Colombian connection? N Engl J a widely disseminated conjugative genta-
their positions within the clade most Med 2008;359:2177-9.
micin resistance (pMG1-like) plasmid and
com monly associated with methicillin- 9. Reyes J, Rincón S, Díaz L, et al. Dis- the emergence of vancomycin resistance
resistant S. aureus hospital-acquired in- semination of methicillin-resistant Staphylo in Enterococcus faecium. J Clin Microbiol
fection in the United States. MBio 2012; coccus aureus USA300 sequence type 8 lin- 2002;40:3326-33.
eage in Latin America. Clin Infect Dis 2009; 16. Arias CA, Panesso D, Singh KV, Rice
2. Limbago BM, Kallen AJ, Zhu W, et al. 49:1861-7.
LB, Murray BE. Cotransfer of antibiotic
Report of the 13th vancomycin-resistant 10. Rodríguez-Noriega E, Seas C, Guzmán- resistance genes and a hylEfm-containing
Staphylococcus aureus isolate from the United Blanco M, et al. Evolution of methicillin- virulence plasmid in Enterococcus faecium.
States. J Clin Microbiol 2014;52:998-1002. resistant Staphylococcus aureus clones in Latin Antimicrob Agents Chemother 2009;53:
3. Saha B, Singh AK, Ghosh A, Bal M. America. Int J Infect Dis 2010;14:e560-6.
Identification and characterization of a 11. Martineau F, Picard FJ, Lansac N, et 17. Barton BM, Harding GP, Zuccarelli
vancomycin-resistant Staphylococcus aureus al. Correlation between the resistance AJ. A general method for detecting and
isolated from Kolkata (South Asia). J Med genotype determined by multiplex PCR sizing large plasmids. Anal Biochem
assays and the antibiotic susceptibility 1995;226:235-40.
4. Azimian A, Havaei SA, Fazeli H, et al. patterns of Staphylococcus aureus and Staph
18. Lozano C, García-Migura L, Aspiroz
Genetic characterization of a vancomycin- ylococcus epidermidis. Antimicrob Agents C, Zarazaga M, Torres C, Aarestrup FM.
resistant Staphylococcus aureus isolate from Chemother 2000;44:231-8.
Expansion of a plasmid classification sys-
the respiratory tract of a patient in a uni- 12. Dutka-Malen S, Evers S, Courvalin P. tem for Gram-positive bacteria and de-
versity hospital in northeastern Iran. J Clin Detection of glycopeptide resistance gen- termination of the diversity of plasmids
otypes and identification to the species in Staphylococcus aureus strains of human,
5. Weigel LM, Clewell DB, Gill SR, et al. level of clinically relevant enterococci by animal, and food origins. Appl Environ
Genetic analysis of a high-level vancomycin- PCR. J Clin Microbiol 1995;33:24-7. [Er ra- Microbiol 2012;78:5948-55.
resistant isolate of Staphylococcus aureus. tum, J Clin Microbiol 1995;33:1434.]
19. Arias CA, Panesso D, McGrath DM, et
13. Woo PC, Leung AS, Leung KW, Yuen al. Genetic basis for in vivo daptomycin
6. Zhu W, Clark N, Patel JB. pSK41-like KY. Identification of slide coagulase posi-
resistance in enterococci. N Engl J Med
plasmid is necessary for Inc18-like vanA tive, tube coagulase negative Staphylococcus 2011;365:892-900.
plasmid transfer from Enterococcus faecalis aureus by 16S ribosomal RNA gene se- 20. Zhu W, Murray PR, Huskins WC, et al.
to Staphylococcus aureus in vitro. Antimicrob quencing. Mol Pathol 2001;54:244-7.
Dissemination of an Enterococcus Inc18-like
Agents Chemother 2013;57:212-9.
14. Performance standards for antimicro-
vanA plasmid associated with vancomycin-
7. DeLeo FR, Otto M, Kreiswirth BN, bial susceptibility testing: twenty-first in-
resistant Staphylococcus aureus. Antimicrob
Chambers HF. Community-associated formational supplement (CLSI document Agents Chemother 2010;54:4314-20.
meticillin-resistant Staphylococcus aureus. no. M100-S21). Wayne, PA: Clinical and 21. O'Brien FG, Coombs GW, Pearman JW,
Laboratory Standards Institute, 2012.
et al. Population dynamics of methicillin-
n engl j med 370;16 nejm.org april
The New England Journal of Medicine
Downloaded from nejm.org at FIOCRUZ on April 24, 2014. For personal use only. No other uses without permission.
Copyright 2014 Massachusetts Medical Society. All rights reserved.

Transferable Vancomycin Resistance in MRSA Lineage
susceptible and -resistant Staphylococcus au Staphylococcus aureus containing the vanA 25. Jiménez JN, Ocampo AM, Vanegas JM,
reus in remote communities. J Anti microb resistance gene. N Engl J Med 2003;348: et al. CC8 MRSA strains harboring SCCmec
type IVc are predominant in Colombian
22. McCarthy AJ, Lindsay JA. The distri-
24. Arthur M, Molinas C, Depardieu F, hospitals. PLoS One 2012;7(6):e38576.
bution of plasmids that carry virulence Courvalin P. Characterization of Tn1546, a 26. Diep BA, Gill SR, Chang RF, et al.
and resistance genes in Staphylococcus au Tn3-related transposon conferring glyco- Complete genome sequence of USA300,
reus is lineage associated. BMC Microbiol peptide resistance by synthesis of depsi- an epidemic clone of community-acquired
peptide peptidoglycan precursors in En ter meticillin-resistant Staphylococcus aureus.
23. Chang S, Sievert DM, Hageman JC, et o coccus faecium BM4147. J Bacteriol 1993; Lancet 2006;367:731-9.
al. Infection with vancomycin-resistant 175:117-27.
Copyright 2014 Massachusetts Medical Society.
Yosemite National Park, California
Brad Warner, M.D.
n engl j med 370;16 nejm.org april
The New England Journal of Medicine
Downloaded from nejm.org at FIOCRUZ on April 24, 2014. For personal use only. No other uses without permission.
Copyright 2014 Massachusetts Medical Society. All rights reserved.
Source: https://www.bio.fiocruz.br/images/stories/pdfs/mpti/2015/transferable-vancomycin-resistance-in-a-community-associated-mrsa-lineage.pdf
Extended spectrum beta-lactamases A. Beta-lactam antibiotics a. d. Mechanism of resistances B. Beta-lactamases a. b. Extended spectrum beta-lactamases (ESBL) c. Non-TEM, non-SBV ESBL d. Inhibitor Resistant TEM (IRT) C. Definition, classification and properties of ESBL D. Epidemiology and risk factors E. Laboratory detection and identification of ESBLs
Felipe Cortés-Salazar1, Josué Omar Suárez Ortíz1, Nancy Mónica Cendejas Trejo1, Juan Manuel Mancilla-Díaz1, Verónica Elsa López-Alonso1, Rodrigo Erick Escartín-Pérez1* 1Neurobiology of Eating Laboratory, Universidad Nacional Autónoma de México, FES Iztacala. Tlalnepantla, México Recibido, mayo 2/2014 Referencia: Cortés-Salazar, F., Suárez Ortíz, J.



