Viagra gibt es mittlerweile nicht nur als Original, sondern auch in Form von Generika. Diese enthalten denselben Wirkstoff Sildenafil. Patienten suchen deshalb nach viagra generika schweiz, um ein günstigeres Präparat zu finden. Unterschiede bestehen oft nur in Verpackung und Preis.
Doi:10.1016/j.procbio.2005.10.012
Process Biochemistry 41 (2006) 930–934
Short communication
Effect of Vitreoscilla hemoglobin gene (vgb) and metabolic inhibitors on
cadmium uptake by the heterologous host Enterobacter aerogenes
Khaled M. Khleifat Muayad M. Abboud, Ahmed H. Al-Mustafa
Mutah University, Department of Biology, P.O. Box (7) Karak, Mutah 61710, Jordan
Received 18 May 2005; received in revised form 29 September 2005; accepted 6 October 2005
The effect of metabolic inhibitor/uncoupler(s) (CCCP and NaN3) and sulfhydryl reagents (dithiothreitol, 2-mercaptoethanol glutathione) on
cadmium uptake was investigated in Enterobacter aerogenes strains. They include a transformed strain bearing the Vitreoscilla hemoglobin gene,vgb as well as control strains that lack this transformed gene. The vgb-harboring strains showed better uptake of cadmium than vgb-lacking strains.
Under low aeration, there was 2-fold enhancement of Cd+2 uptake in vgb-harboring strains compared with 1.6-fold enhancement under highaeration. The CCCP caused 36, 40 and 58% inhibition in cadmium uptake of parental, pUC9-harboring and VHb-expressing cells, respectively.
Similarly, the sodium azide exerted 44, 38 and 55% inhibition in Cd+2 uptake of parental, pUC9-harboring and VHb-expressing cells, respectively.
Less extensive inhibition of Cd+2 uptake in the range of 11–39% was observed with sulfhydryl reagents.
# 2005 Elsevier Ltd. All rights reserved.
Keywords: Bacterial hemoglobin; Cadmium uptake; Enterobacter aerogenes
stranded breakage of cellular DNA or may interfere with therespiration and growth of bacteria . The mechanism of Cd+2
Most heavy metals are toxic to microorganisms, particularly
resistance in Gram-negative bacteria is primarily based on the
at high concentration, due to the complexing of these metals with
action of an efflux system called RND that have diverse functions
certain compounds inside the cell Heavy metals must first
like resistance/nodulation of legumes/cell division . This
enter the bacterial cell before they produce their toxic effects.
active transporter is proton-driven export system .
Generally, there are two major mechanisms for bacterial metal
Despite the detection of such metal resistance system in
uptake . The first is ATP independent, non-specific, fast and
several Gram-negative bacteria little is known on the
govern by the chemiosmotic gradient across the cell membrane.
cadmium resistance system in Enterobacter aerogenes.
The second is ATP dependent, slower and more metal-specific.
Furthermore, no previous work have been reported regarding
The natural and industrial processes may increasingly
the implication of bacterial hemoglobin on general metal
release heavy metals to microbial habitats. Therefore, microbes
resistance in bacteria.
have adapted different mechanisms to tolerate the stress of
The presence of hemoglobin protein in prokaryotic
heavy metals These mechanisms mainly include the
organisms was first recognized in the Gram-negative bacterium
metal efflux outside the cell or a complexing with organic
Vitreoscilla This Vitreoscilla hemoglobin is the most
compounds inside the cell or the chemical reduction of heavy
widely studied bacterial hemoglobin, including its potential use
metals to less toxic state
in biotechnological applications . Enhanced biosynthesis of
In Gram-negative bacteria such as E. coli, the active uptake of
VHb is mediated at the transcriptional level by an oxygen-
cadmium is coupled with the Zn+2 transport system . Once
sensitive promoter, which is turned on under hypoxic
inside the cell, cadmium may become toxic either through single-
conditions in the native and recombinant host of E. coli . The expression of VHb hemoglobin may result in theenhancement of cell density, oxidative metabolism, bioreme-diation and engineered product formation especially under
Abbreviations: VHb, Vitreoscilla hemoglobin; vgb, hemoglobin gene;
oxygen-limited conditions. A particular application of VHb
CCCP, carbonyl cyanide chlorophenyl hydrazone
expression involved the increased production of a-amylase
* Corresponding author.
E-mail address:
[email protected] (K.M. Khleifat).
, the enhanced degradation of toxic wastes such as 2,4-DNT
1359-5113/$ – see front matter # 2005 Elsevier Ltd. All rights reserved.
doi:10.1016/j.procbio.2005.10.012
K.M. Khleifat et al. / Process Biochemistry 41 (2006) 930–934
and benzoic acid degradation by Pseudomonads . It is
azide (NaN3), dithiothreitol, and glutathione that were used at 10 mM con-
believed that the putative function of VHb is to trap oxygen and
feeds it to a membrane terminal oxidase The general effect
2.4. Measurement of cadmium biosorption, adsorption and uptake
of VHb is based on the fact that a bacterial cells engineered withthe gene (vgb) encoding Vitreoscilla hemoglobin (VHb)
The biosorption of Cd+2 was carried out according to the method of Scott
typically produce more protein than vgb-lacking cells
and Kanazawa and Mori with slight modifications. Cultures of the three
strains were grown in LB medium at 37 8C using shaking incubator under
The present work investigated the effect of vgb on Cd+2
agitation rate of 150 rev/min. Cells were harvested by centrifugation at8000 rev/min for 20 min. The supernatant was then discarded and the cells
biosorption, uptake and adsorption by E. aerogenes trans-
washed twice with 0.85% saline. Harvested biomass (50 mg) was resuspended
formed with vgb gene. Such effect was compared with matched
in 12 ml LB containing of 0, 25, 50, 75, 100 150 and 200 mg l 1 Cd(NO3)2 for
strains lacking vgb and conducted at both high as well as low
1 h. These solutions were filtered by Millipore filter (0.45 mM in pore size), to
level of aerations.
separate the biomass from the filtrate. Subsequently, the biomass was washedeither with distilled water or with 0.01N-CH3COONH4 solutions at pH 4.0according to the method of Kanazawa and Mori After washing, the biomass
2. Materials and methods
was reweighed in the dry state and decomposed by 1% nitric acid for 24 h. Cd+2content for the latest solution was determined by using a flame atomic
2.1. Growth conditions
absorption spectrophotometer [spect AA 800 (Varian) with deuterium lampand background correction]. The water-washed biomass was used for measur-
The host bacterium Enterobacter aerogenes was obtained from Microbiol-
ing the Cd+2 biosorption, while the CH3COONH4 washed biomass was used for
ogy Laboratory at the Department of Biology, Mutah University (Jordan). Its
the Cd+2 uptake. The Cd+2 adsorption was then determined by subtracting the
identity has been confirmed previously . Cells were transformed with two
amount of Cd+2 uptake from the amount of Cd+2 biosorption.
different plasmids and grown on either LB or LB-added ampicillin (AP) media.
The first plasmid is called pUC8:16, which contains the vgb gene located on
2.5. Examining the effect of agitations rate on the Cd+2 uptake,
1.4 kb fragment This plasmid was obtained from the Biology Department,
adsorption and biosorption
Illinois Institute of Technology (Chicago, IL, USA). The second plasmid(pUC9) similar to pUC8:16 but lacks the bacterial hemoglobin gene, vgb
The LB media was used as mentioned previously to grow bacterial cells at
This plasmid was provided as a gift by Professor Khaled Al-Tarawneh
37 8C under two agitation rates 250 and 75 rev/min. These agitation rates are
from the Biology Department, Mutah University (Jordan). The resulting two
referred to as high and low aeration, respectively . To examine the effect
strains were denoted Enterobacter:pUC8:16 and Enterobacter:pUC9, respec-
of aeration on the uptake of Cd+2, an exponentially growing cells were first
tively. Both transformants were checked for the harbored plasmids by using
collected by centrifugation and then resuspended in Cd+2 solutions as described
agarose gel electrophoresis of plasmid minipreparations. The vgb recombinants
were easily recognizable because of their large accumulation of VHb and theircharacteristic carbon monoxide difference spectra .
2.2. Determining the toxicity effect of Cd on bacterial growth
The bacterial growth medium composed of LB supplemented with 0, 50,
3.1. Effect of agitation rate on bacterial mass, Cd uptake,
100, 150 ppm cadmium nitrate. Medium for the growth of transformed strains
biosorption and adsorption
additionally contained ampicillin at 50 mg/ml concentration. The cells wereincubated in a GFL Model 3032 shaker (Burgwedel, Germany) at 37 8C using
The Enterobacter strains yielded more cell biomass under
normal agitation rate of 150 rev/min. Samples were taken at intervals todetermine the OD at 600 nm. The growth curves based on OD
high aeration than under low aeration (Moreover,
were made, and an average of the first five OD
these strains showed elevated uptake of Cd+2, which was
600 measurements within the
stationary phase was taken as a final reading. The same time interval was taken
initially, proportional to the concentration of Cd+2 being used in
into consideration for the three Enterobacter strains.
the medium (). The cadmium uptake exhibitedMichaelis–Menten saturation kinetics where the maximum
2.3. Testing the inhibitors/uncoupler effects
saturation point in vgb-bearing strain was higher than the valueobserved in vgb-lacking strains.
Exponential Enterobacter strain cells were preincubated for 20 min with
various metabolic inhibitors at 20 8C in a standard assay medium. These
Variable rates of Cd+2 uptake were obtained when
inhibitors include carbonyl cyanide chlorophenyl hydrazone (CCCP), sodium
Enterobacter cells were grown at different aeration conditions
Table 1Aeration dependence of Cd uptake, adsorption and biosorption by Enterobacter aerogenes (parental, E:pUC9 and E:pUC8:16) strains and their cell biomass
Values are the average of three individual experiments; standard deviations are in parenthesis.
K.M. Khleifat et al. / Process Biochemistry 41 (2006) 930–934
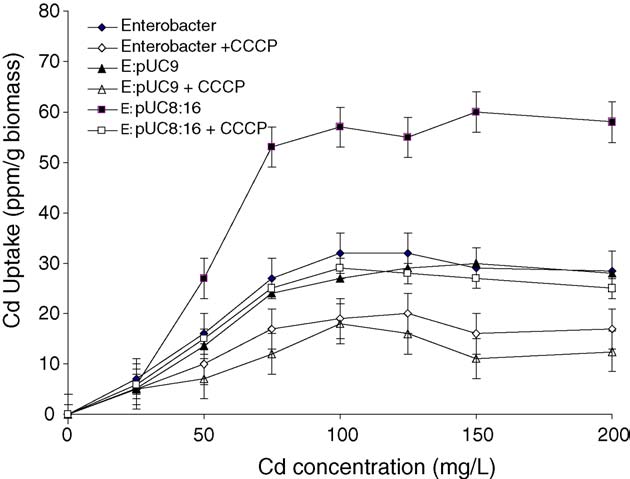
Fig. 1. Effect of Cd+2 concentration in the medium on Cd+2 uptake byEnterobacter aerogenes strains (parental, E:pUC9 and E:pUC8:16). The uptake
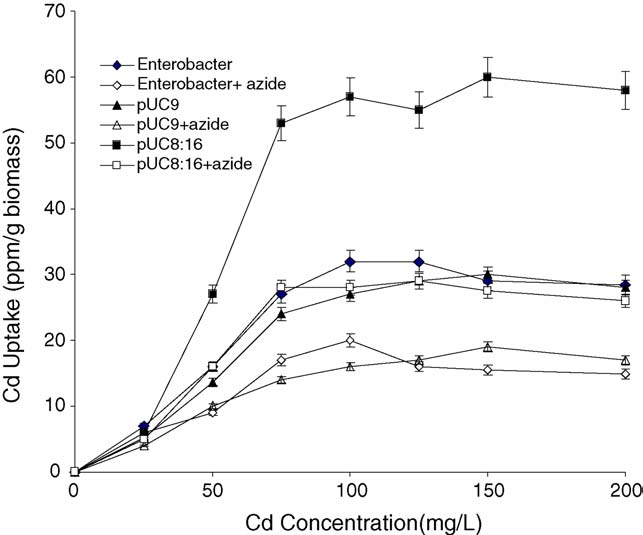
Fig. 3. Effect of 10 mM NaN
assay was performed at 37 8C as described in Section Each point represents
3 on the Cd+2 uptake by Enterobacter aerogenes
strains (parental, E:pUC9 and E:pUC8:16). Uptake assays was performed at
the mean � S.D. (n = 3).
37 8C as described in Section Each point represents the mean � S.D. (n = 3).
(Under low aerations (75 rev/min), the rate of Cd+2uptake in the vgb-bearing strain was almost twice the rate
By comparing the initial saturation point of inhibition, the
detected in the two vgb-lacking strains. Also, the vgb-bearing
Cd+2 uptake in vgb-bearing strain scored 58% inhibition in
strain showed a decrease in cadmium adsorption and increase
presence of CCCP () and 55% in presence of sodium
in biosorption. Under high aerations (250 rev/min), though
azide, respectively (A similar effect by CCCP on VHb-
the Cd+2 uptake in vgb-bearing strain was much higher
lacking strains caused nearly 36% inhibition of Cd+2 uptake in
(56.2 ppm/g) than that detected under low aerations, but the
parented cells and 40% in pUC9 strain, respectively. Moreover,
difference in Cd+2 uptake between the vgb-bearing and the
sodium azide caused 44% inhibition of the uptake in parental
vgb-lacking strains was only 1.6-fold. At similar conditions,
and 38% inhibition in pUC9 harbouring cells, respectively.
all strains exhibited general rise in Cd+2 adsorption and
shows further inhibition of Cd+2 uptake ranged
between 11–39%, exerted by the sulfhydryl reagents 2-mercaptoethanol, dithiothreitol and glutathione. However, such
3.2. Effect of inhibitors/uncouplers on Cd+2 uptake
The addition of CCCP or sodium azide in vitro produced
marked inhibition effect on Cd+2 uptake Thisinhibition was detected in all Enterobacter strains but its degreewas variable.
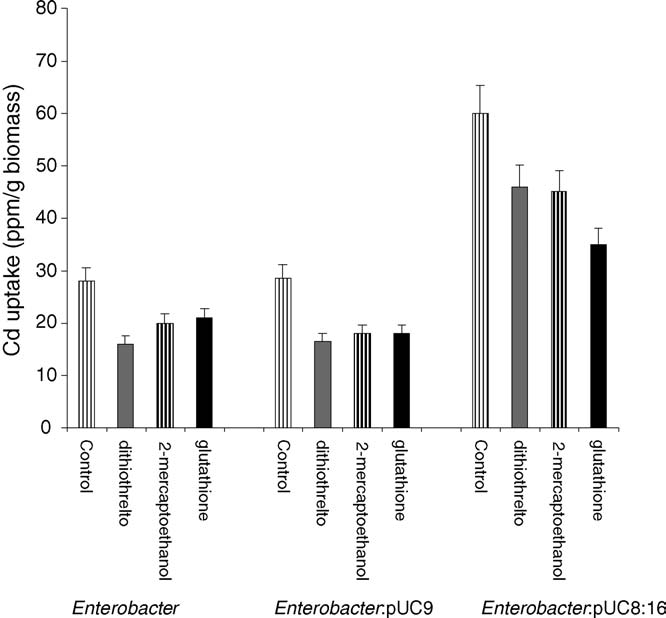
Fig. 4. Effect of 10 mM sulfhydryl reagents (2-mercaptoethanol, dithiothreitol
Fig. 2. Effect of 10 mM CCCP on the Cd+2 uptake by Enterobacter aerogenes
and glutathione) on the Cd+2 uptake by Enterobacter aerogenes strains (par-
strains (parental, E:pUC9 and E:pUC8:16). Uptake assay was performed at
ental, E:pUC9 and E:pUC8:16). Uptake assay was performed at 37 8C as
37 8C as described in Section . Each point represents the mean � S.D. (n = 3).
described in Section Results represent the mean � S.D. (n = 3).
K.M. Khleifat et al. / Process Biochemistry 41 (2006) 930–934
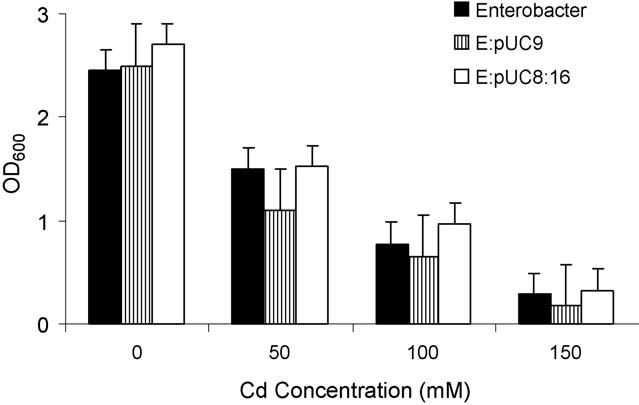
the effect of cadmium on bacterial growth suggest that VHb-expressing strains endure better resistance to Cd+2 toxicity thanthe two control strains.
In E. coli, the MIC for cadmium is 0.5 mm, which is
considered with platinum as one of the most toxic heavy metalsThe bacterial resistance to cadmium in bacteria seems to bedependent mainly on the mechanism of metal efflux. In Gram-negative bacteria, the cadmium is primarily detoxified by thezinc exporter RND-protein driven pump . It is possible thatthe expression of VHb in the transformed Enterobacter strainhave inducting effect on the RND system of cadmium exporter.
The important role played by bacterial hemoglobin in
metabolic activities makes this protein a suitable candidate
Fig. 5. Toxicity of Cd+2 against Enterobacter aerogenes strains (parental,
for controlling heavy metal detoxification. Its known capability
E:pUC9 and E:pUC8:16) cultured in the LB buffered medium. Cd+2 concen-
to enhance the production of cellular proteins may either
trations used were 0, 50, 100 and 150 ppm. The growth curves based on OD600
contribute in a general or selective manner to the Cd+2
measurements were made, then the average of the first five OD600 measure-ments of the stationary phase was taken as a final reading after 24 h. Results
transporter protein activity.
represent the mean � S.D. (n = 3).
The enhancement in Cd+2 uptake by the VHb-bearing strain
under low aeration is in keeping with the function of VHb as an
inhibition was less extensive than the inhibition detected with
oxygen-trap protein. The oxy-VHb may act positively on some
CCCP and sodium azide.
key redox component of the cell, which could be a sensor, aregulator or an allosteric site of respiratory protein .
3.3. Toxicity effect of Cd on bacterial growth
This may explain the high sensitivity of Cd+2 uptake to theinhibition of uncouplers CCCP and sodium azide observed
When cadmium was added in vivo to different Enterobacter
particularly in the vgb-bearing transformed strain. The slight
strains, it became toxic particularly at high concentrations
inhibition of Cd+2 uptake by thiol compounds might be the
(). This toxicity effect of Cd+2 was expressed by the rate
outcome of an extracellular complexation between the Cd+2
of bacterial growth suppression relative to a control of bacterial
and the thiol compound. A Cd–S complex has been
growth without cadmium.
demonstrated in Klebsiella pneumoniae and Saccharomyces
The vgb-bearing cells persistently gave a better growth rate
cerevisiae as a mechanism of Cd+2 detoxification
than the other two vgb-lacking control strains, when they were
In conclusion, evidences are presented in this work for an
subjected to various concentrations of cadmium.
active transport system of Cd+2 in Enterobacter aerogenes. Thisevidence is indicated by the Michaelis–Menten saturation
pattern of Cd+2 uptake, the decrease of this transport rate byinhibitors of cellular energy and thiol compounds, which was
So far, few genetically engineered methods have been
then particularly relieved by VHb expression. To our knowl-
manipulated to improve the bacterial accumulation of
edge, this is the first report on the use of engineering bacterial
environmental toxic metals These methods mainly
hemoglobin for improving metal accumulation. The impact of
involve the fusion of external metal-binding adapters such as
this improvement on bacterial bioremediation of toxic heavy
metallothionein or short polypeptide to the bacterial cell
metals will await further research work.
surface. The short polypeptide proved to have better advantageover the metallothionein adapter but still both
methods in general carry certain limitations. The instability ofadapter fusion, the lack of selective metal binding, the decrease
We would like to thank the science research deanship at
in affinity of metal binding and the reduction in metal tolerance,
Mutah University for supporting this research.
are some of the difficulties that potentially may encounter.
To avoid all these difficulties, we have introduced a new
genetically engineered method for the bacterial transport of
heavy metals from surrounding environment. This method is
[1] Nies DH, Silver S. Plasmid-determined inducible efflux is responsible for
based on the transformation of Vitreoscilla hemoglobin gene-
resistance to cadmium, zinc, and cobalt in Alcaligenes eutrophus. J
containing plasmid, pUC8:16 into the bacteria Enter-
obacter aerogenes. We have shown that the transformed vgb-
[2] Nies DH, Silver S. Ion efflux systems involved in bacterial metal resis-
bearing Enterobacter strain with Vitreoscilla hemoglobin gene,
tances. J Indust Microbiol 1995;14(2):186–99.
vgb, is capable of improving the cadmium uptake. To the
[3] Nies DH. The cobalt, zinc, and cadmium efflux system CzcABC from
Alcaligenes eutrophus functions as a cation–proton antiporter in Escher-
contrary, control strains of parental Enterobacter or trans-
ichia coli. J Bacteriol 1995;177(10):2707–12.
formed strains that lack vgb, have failed to achieve a similar
[4] Gadd GM, Griffiths AJ. Microorganisms and heavy metal toxicity. Microb
improvement in Cd+2 uptake. Furthermore, data obtained from
K.M. Khleifat et al. / Process Biochemistry 41 (2006) 930–934
[5] Khazeli MB, Mitra RS. Cadmium-binding component in Escherichia coli
[21] Khosla C, Curtis JE, DeModena JA, Rinas U, Bailey JE. Expression of
during accommodation to low levels of this ion. Appl Environ Microbiol
intracellular hemoglobin improves protein synthesis in oxygen-limited
Escherichia coli. Bio/Technology 1990;8:849–53.
[6] Higham DP, Sadler PJ, Scawen MD. Cadmium resistant Pseudomonas
[22] Liu SC, Webster DA, Stark BC. Cloning and expression of the Vitreoscilla
hemoglobin gene in Pseudomonas: effects on cell growth. Appl Microbiol
[7] Kanazawa S, Mori K. Isolation of cadmium-resistant bacteria & their
[23] Dikshit KL, Webster DA. Cloning, characterization and expression of the
resistance mechanisms: Part 11. Cadmium biosorption by Cd-resistant and
bacterial hemoglobin gene from Vitreoscilla in Escherichia coli. Gene
sensitive bacteria. Soil Sci Plant Nutrit 1996;42(4):731–6.
[8] Hao Z, Chen S, Wilson DB. Cloning, expression and characterization of
[24] Scott JA. Sites of cadmium uptake in bacteria used for biosorption. Appl
cadmium and manganese uptake genes from Lactobacillus plantarum.
Environ Microbiol 1990;33:221–5.
Appl Environ Microbiol 1999;65(11):4746–52.
[25] Bae W, Chen W, Mulchandani A, Mehra RK. Enhanced bioaccumulation
[9] Mitra RS, Bernstein LA. Single-strand breakage in DNA of Escherichia
of heavy metals by bacterial cells displaying synthetic phytochelatins.
coli exposed to Cd+2. J Bacteriol 1978;133:75–80.
Biotechnol Bioeng 2000;70:518–24.
[10] Holmes JD, Richardson DJ, Saed S, Evans-Gowing R, Russell DA, Sodeau
[26] Bae W, Mehra RK, Mulchandani A, Chen W. Genetic engineering of
JR. Cadmium-specific formation of metal sulfide ‘Q particles' by Kleb-
Escherichia coli for enhanced uptake and bioaccumulation of mercury.
siella pneumonia. Microbiology 1997;143:2521–30.
Appl Environ Microbiol 2001;67:5335–8.
[11] Sair MH, Tarn R, Reizer A, Reizer J. Two novel families of bacterial
[27] Li Y, Cockburn W, Kilpatrick J, Whitelam GC. Cytoplasmic expression of
membrane proteins concerned with nodulation, cell division and transport.
a soluble synthetic mammalian metallothionein-alpha domain in Escher-
Mol Microbiol 1994;11:841–7.
ichia coli enhanced tolerance and accumulation of cadmium. Mol Bio/
[12] Rensing C, Pribyl T, Nies DH. New functions for the three subunits of the
czcCBA cation–proton–antiporter. J Bacteriol 1997;22:6871–9.
[28] Mejare M, Bulow L. Metal-binding proteins and peptides in bioremedia-
[13] Horitsu H, Yamamoto K, Wachi S, Fukuchi A. Plasmid-determined
tion and phytoremediation of heavy metals. Trends Biotechnol 2001;19:
cadmium resistance in Pseudomonas putida GAM-I isolated from soil.
J Bacteriol 1986;165:334–5.
[29] Pazirandeh M, Chrisey LA, Mauro JM, Campbell JR, Gaber BP. Expres-
[14] Wakabayashi S, Matsubara H, Webster DA. Primary sequence of a dimeric
sion of the Neurospora crassa metallothionein gene in Escherichia coli
bacterial hemoglobin from Vitreoscilla. Nature 1986;322:481–3 (Lon-
and its effect on heavy metal uptake. Appl Microbiol Biotechnol 1995;43:
[15] Pringshem EG. The Vitreoscillaceae: a family of colorless, gliding,
[30] Sousa C, Cebolla A, de Lorenzo V. Enhanced metallo-adsorption of
filamentous organisms. J Gen Microbiol 1951;5:124–49.
bacterial cells displaying poly-His peptides. Nature Biotechnol 1996;
[16] Ramandeep H, Wang KW, Raje M, Kim KJ, Stark BC, Dikshit KL,
Webster DA. Vitreoscilla hemoglobin: intracellular localization and bind-
[31] Sousa C, Kotrba P, Ruml T, Cebolla A, de Lorenzo V. Metalloadsorption
ing to membranes. J Biol Chem 2001;276(27):24781–9.
by Escherichia coli cells displaying yeast and mammalian metallothio-
[17] Joshi M, Dikshit KL. Oxygen dependent regulation of Vitreoscilla globin
neins anchored to the outer membrane protein LamB. J Bacteriol 1998;
gene: evidence for positive regulation by FNR. Biochem Biophys Res
[32] Sauge-Merle S, Cuine S, Carrier P, LecomtePradines C, Luu DT, Peltier G.
[18] Kallio PT, Kim DJ, Tsai PS, Bailley JE. Intracellular expression
Enhanced toxic metal accumulation in engineered bacterial cells expres-
of Vitreoscilla hemoglobin alters Escherichia coli energy metabo-
sing Arabidopsis thaliana phytochelatin synthase. Appl Environ Micro-
lism under oxygen limited condition. Eur J Biochem 1994;219:
[33] Kallio PT, Bailey JE. Intracellular expression of Vitreoscilla hemoglobin
[19] Khleifat K, Abboud MM. Correlation between bacterial hemoglobin gene
(VHb) enhances total protein secretion and improves the production of a-
(vgb) and aeration: their effect on the growth and a-amylase activity
amylase and neutral protease in Bacillus subtilis. Biotechnol Prog 1996;
in transformed Enterobacter aerogenes. J Appl Microbiol 2003;94:
[34] Aiking H, Grover H, Vant' Reit J. Detoxification of mercury, cadmium and
[20] Nasr MA, Hwany KW, Akbas M, Webster DA, Stark BC. Effects of culture
lead in Klebsiella aerogenes NCTC 418 growing in continuous culture.
conditions on enhancement of 2-4-dinitrotolune degradation by Burkhol-
Appl Environ Microbiol Environ Microbiol 1985;50:1262–7.
deria engineered with the Vitreoscilla hemoglobin gene. Biotechnol Prog
[35] Gharieb MM, Gadd GM. Role of glutathione in detoxification of metal(-
loid)s by Saccharomyces cerevisiae. BioMetals 2004;17:83–188.
Source: http://e-print.mutah.edu.jo/index.php/document-repository/doc_download/4-effect-of-vitreoscilla-hemoglobin-gene-vgb-and-metabolic-inhibitors-on.html
J_ID: CHI Customer A_ID: 08-0027 Cadmus Art: CHI20564 Date: 23-MAY-08 Stage: I CHIRALITY 00:000–000 (2008) Use of Large-Scale Chromatography in the Preparation of Armodafinil WILLY HAUCK,1 PHILIPPE ADAM,2 CHRISTELLE BOBIER,2* AND NELSON LANDMESSER3 1Novasep Inc., Boothwyn, Pennsylvania 2Novasep SAS, Pompey, France 3Cephalon Inc., West Chester, Pennsylvania Armodafinil, the (R)-enantiomer of modafinil, is a medication used to
power, performance, price Erectile dysfunction Erectile dysfunction is a common condition affecting over 50% of men over the age of 40, at some time1. How many of your patients are hiding in their sheds or workshops, afraid or too embarrassed to discuss their erectile difficulties and not receiving treatment?Men need good tools. So let's get your male patients out



